História evolutiva da vida
A história evolutiva da vida na Terra traça os processos pelos quais organismos vivos e fósseis evoluíram. A Terra se formou há cerca de 4,5 bilhões de anos (abreviado como Ga, para gigaannum) e as evidências sugerem que a vida surgiu antes de 3,7 Ga.[1][2][3] (Embora haja alguma evidência de vida tão cedo quanto 4,1 a 4,28 Ga, ela permanece controversa devido à possível formação não biológica dos fósseis alegados.[1][4][5][6]) As semelhanças entre todas as espécies atuais conhecidas indicam que elas divergiram ao longo do processo de evolução de um ancestral comum.[7] Aproximadamente 1 trilhão de espécies vivem atualmente na Terra,[8] das quais apenas 1,75-1,8 milhões foram nomeadas[9][10] e 1,8 milhões documentadas em um banco de dados central.[11] Essas espécies vivas atualmente representam menos de um por cento de todas as espécies que já viveram na Terra.[12][13]
As evidências da existência de vida mais antiga vem de amostras de carbono biogênicos[2][3] e fósseis de estromatólito[14] descobertos 3,7 bilhões de anos, as rochas metassedimentares descobertas no oeste da Groenlândia. Em 2015, possíveis "restos de vida biótica" foram encontrados em rochas de 4,1 bilhões de anos na Austrália Ocidental.[15][16] Em março de 2017, a evidência putativa de possivelmente as formas mais antigas de vida na Terra foi relatada na forma de micro-organismos fossilizados descobertos em precipitados de fontes hidrotermais no Cinturão Nuvvuagittuq de Quebec, Canadá, que podem ter vivido há 4,28 bilhões de anos, não muito depois da formação dos oceanos, 4,4 bilhões de anos atrás, e não muito depois da formação da Terra, 4,54 bilhões de anos atrás.[17][18]
Biofilmes de bactérias e archaea coexistentes foram a forma de vida dominante no início do Arqueano e pensa-se que muitos dos principais passos nos primórdios da evolução tiveram lugar dentro deles.[19] A evolução de fotossíntese com oxigênio, há cerca de 3,5 bilhões de anos, eventualmente levou à oxigenação da atmosfera, começando por volta de há 2 400 milhões de anos.[20] Enquanto que células eucariotas podem ter estado presentes anteriormente, a sua evolução foi acelerada quando começaram a usar o oxigénio no seu metabolismo. A evidência mais antiga de eucariotas complexos com organelos, data de 1,85 bilhões de anos. Mais tarde, por volta de há 1 700 milhões de anos, começaram a aparecer organismos multicelulares, com células diferenciadas a realizar funções especializadas.[21]
As primeiras plantas terrestres datam de há cerca de 450 milhões de anos,[22] apesar de evidências sugerirem que algas formaram-se em terra tão cedo com há 1,2 bilhões de anos. Plantas terrestres foram tão bem sucedidas que se pensa que elas contribuíram para a extinção do Devoniano.[23] Os animais invertebrados apareceram durante o Ediacarano,[24] enquanto que os vertebrados surgiram há cerca de 525 milhões de anos, durante a explosão do Cambriano.[25]
Durante o Permiano, os sinápsidos, incluindo os ancestrais de mamíferos, dominaram a terra,[26] mas a porém com a extinção do Permiano-Triássico há 251 milhões de anos, este fato esteve perto de dizimar toda a vida complexa.[27] Durante a recuperação desta catástrofe, os Archosauria tornaram-se os vertebrados terrestres mais abundantes, substituindo os therapsida em meados do Triássico.[28] Um grupo de archosauria viveram quando os dinossauros, dominaram o Jurássico e Cretácico,[29] enquanto os ancestrais dos mamíferos sobreviviam como pequenos insectívoros.[30] Depois da extinção Cretáceo-Paleogeno há 65 milhões de anos ter morto os dinossauros não-avianos[31] os mamíferos aumentaram rapidamente em tamanho e diversidade.[32] Tal extinção em massa pode ter acelerado a evolução ao fornecer oportunidades para novos grupos de organismos de diversificar.[33]
Evidências fósseis indicam que as plantas com flor apareceram e rapidamente diversificaram no princípio do Cretácico, entre há 130 milhões a 90 milhões de anos, provavelmente pela coevolução com insectos polinizadores. Plantas com flores e fitoplâncton marinho são ainda os produtores de matéria orgânica dominantes. Insectos sociais apareceram por volta da mesma altura que as plantas com flor. Apesar de ocuparem apenas uma pequena parte da "árvore da vida" dos insectos, agora formam cerca de metade da massa total dos insetos. Os humanos evoluíram a partir de uma linhagem com diferentes espécies de hominídeos cujos fósseis mais antigos datam de há mais de 6 milhões de anos. Apesar dos membros mais antigos desta linhagem terem cérebros do tamanho semelhante ao de um chimpanzé, há sinais de um aumento constante do tamanho do cérebro após 3 milhões de anos.
História inicial da Terra
editarplantas mais antigos
Os mais antigos fragmentos de meteorito encontrados na Terra têm cerca de 4,54 bilhões de anos de idade; isto, junto principalmente com a datação de depósitos de chumbo antigos, colocou a estimada idade da Terra por volta deste tempo.[36] A Lua tem a mesma composição da crosta terrestre mas não contém um núcleo planetário rico em ferro como o da Terra. Muitos cientistas pensam que cerca de apenas 40 milhões de anos depois um planetoide atingiu a Terra, lançando para o espaço material da crosta que acabou por formar a Lua. Uma outra hipótese é que a Terra e a Lua começaram a coalescer ao mesmo tempo, mas a Terra, tendo uma gravidade muito mais forte, atraiu quase todas as partículas de ferro na área.[37]
Até recentemente, as rochas mais antigas encontradas na Terra foram estimadas possuírem cerca de 3,8 bilhões de anos de idade,[36] levando cientistas a acreditar durante décadas que a superfície da Terra estava fundida até essa altura. De acordo com isto, nomearam esta parte da história da Terra de éon Hadeano, significando "infernal".[38] No entanto, análises de zircons formados entre 4,0 a 4,4 bilhões de anos indicam que a crosta solidificou cerca de 100 milhões de anos depois da formação do planeta e que o planeta rapidamente adquiriu oceanos e uma atmosfera, que podem ter sido capazes de suportar vida.[39]
Evidências recolhidas da lua indicam que a partir de 4,0 a 3,8 bilhões de anos sofreu um Intenso bombardeio tardio por detritos que sobraram da formação do Sistema Solar, e a Terra deveria ter sofrido um bombardeamento ainda mais intenso devido à sua maior gravidade.[38][40] Apesar de não haver evidência direta das condições na Terra há 4,0 - 3,8 bilhões de anos, não há razões para pensar que a Terra não foi afectada por este intenso bombardeamento tardio.[41] O evento poderá ter removido qualquer atmosfera e oceanos anteriores; neste caso gases e água resultantes de impactos de cometa podem ter contribuído para a sua substituição, apesar de que a desgaseificação vulcânica na Terra teria contribuído pelo menos metade.[42] No entanto, se a vida microbiana subterrânea tivesse evoluído neste ponto, ela teria sobrevivido ao bombardeio.[43]
Evidências mais antigas da vida na Terra
editarOs organismos mais antigos que já foram identificados eram diminutos e com relativamente poucas características, e os seus fósseis parecem-se com pequenos bastonetes, que são difíceis de diferenciar de estruturas que surgem através de processos físicos abióticos. A evidência mais antiga indisputável da vida na Terra, interpretada como bactérias fossilizadas, data de há 3 mil milhões de anos.[44] Outras descobertas em rochas datadas em cerca de 2,5 mil milhões de anos têm sido também interpretadas como bactérias,[45] com evidências geoquímicas aparentemente mostrando a presença de vida há 3,8 mil milhões de vida.[46] Contudo estas análises foram escrutinadas cuidadosamente, e foram encontrados processos não-biológicos que poderiam produzir todos estes "sinais de vida" que foram relatados.[47][48] Embora isto não prove que as estruturas encontradas tenham uma origem não-biológica, elas não podem ser tomadas como evidências claras para a presença de vida. Assinaturas geoquímicas de rochas depositadas há 3,4 mil milhões de anos foram interpretadas como evidências de vida,[44][49] embora estas afirmações não tenham sido examinadas pormenorizadamente por críticos.
Evidências de microorganismos fossilizados considerados entre 3,77 bilhões a 4,28 bilhões de anos foram encontradas no Cinturão de Rochas Verdes Nuvvuagittuq em Quebec, Canadá,[17] embora as evidências sejam contestadas como inconclusivas.[50]
Origem da vida na Terra
editar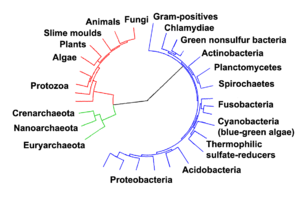
Todos os organismos vivos na Terra devem ter um único ancestral universal, porque seria virtualmente impossível para duas ou mais linhagens separadas serem capazes de desenvolver independentemente os muitos mecanismos bioquímicos complexos comuns a todos os organismos vivos.[52][53]
Vida "semeada" de outro lugar
editarA ideia de que a vida na Terra foi "semeada" de outras partes do universo data pelo menos do século V.[54] No século XX foi proposto pelo químico físico Svante Arrhenius,[55] pelos astrónomos Fred Hoyle e Chandra Wickramasinghe,[56] e pelo biólogo molecular Francis Crick e pelo químico Leslie Orgel.[57] Há três versões principais para a hipótese de "sementeira de outro local": vindo de outro lugar do nosso Sistema Solar via fragmentos lançados para o espaço por um impacto de um meteoro de grandes dimensões, sendo a única fonte credível Marte;[58] por visitantes extra-terrestres, possivelmente por contaminação acidental com microorganismos que trouxeram com eles;[57] e de fora do Sistema Solar mas por meios naturais.[55][58] Experimentos em órbitas baixas da Terra, como EXOSTACK, mostraram que alguns esporos de microrganismos podem sobreviver ao choque de serem catapultados para o espaço e alguns podem sobreviver à exposição à radiação do espaço exterior por pelo menos 5,7 anos (duração do experimento).[59][60] Os cientistas estão divididos quanto à probabilidade de que a vida se originou independentemente em Marte,[61] ou em outros planetas em nossa galáxia.[58]
Aparição independente na Terra
editarA vida na Terra é baseada em carbono e água. O carbono fornece estruturas estáveis para produtos químicos complexos e pode ser facilmente extraído do meio ambiente, especialmente do dióxido de carbono. Não há nenhum outro elemento químico cujas propriedades sejam semelhantes o suficiente às do carbono para ser chamado de análogo; silício, o elemento diretamente abaixo do carbono na tabela periódica, não forma muitas moléculas estáveis complexas, e porque a maioria de seus compostos são insolúveis em água e porque o dióxido de silício é um sólido duro e abrasivo em contraste com o dióxido de carbono em temperaturas associadas com seres vivos, seria mais difícil para os organismos extrair. Os elementos boro e fósforo possuem químicas mais complexas, mas apresentam outras limitações em relação ao carbono. A água é um excelente solvente e tem duas outras propriedades úteis: o fato de que o gelo flutua permite que os organismos aquáticos sobrevivam sob ele no inverno; e suas moléculas têm extremidades eletricamente negativas e positivas, o que permite que ele forme uma gama mais ampla de compostos do que outros solventes. Outros bons solventes, como a amônia, são líquidos apenas em temperaturas tão baixas que as reações químicas podem ser muito lentas para sustentar a vida e não têm as outras vantagens da água.[62] Organismos baseados em bioquímica alternativa podem, entretanto, ser possíveis em outros planetas.[63]
A pesquisa sobre como a vida pode ter surgido de substâncias químicas não vivas concentra-se em três pontos de partida possíveis: autor-replicação, a capacidade de um organismo de produzir descendentes muito semelhantes a ele; metabolismo, sua capacidade de se alimentar e se reparar; e membranas celulares externas, que permitem a entrada de alimentos e a saída de resíduos, mas excluem substâncias indesejadas.[64] A pesquisa sobre abiogênese ainda tem um longo caminho a percorrer, pois as abordagens teórica e empírica estão apenas começando a fazer contato uma com a outra.[65][66]
Primeira replicação: mundo de RNA
editarMesmo os membros mais simples dos três domínios modernos da vida usam o DNA para registrar suas "receitas" e um conjunto complexo de moléculas de RNA e proteínas para "ler" essas instruções e usá-las para crescimento, manutenção e autor-replicação. A descoberta de que algumas moléculas de RNA podem catalisar sua própria replicação e a construção de proteínas levou à hipótese de formas de vida anteriores baseadas inteiramente em RNA.[67] Essas ribozimas poderiam ter formado um mundo de RNA no qual havia indivíduos, mas nenhuma espécie, pois mutações e transferências gênicas horizontais significariam que a prole em cada geração provavelmente teria genomas diferentes daqueles com os quais seus pais começaram.[68] Mais tarde, o RNA teria sido substituído pelo DNA, que é mais estável, e portanto, pode construir genomas mais longos, expandindo a gama de capacidades que um único organismo pode ter.[68][69][70] As ribozimas permanecem como os principais componentes dos ribossomos, as "fábricas de proteínas" das células modernas.[71] As evidências sugerem as primeiras moléculas de RNA formadas na Terra antes de 4,17 Ga.[72]
Embora moléculas curtas de RNA auto-replicantes tenham sido artificialmente produzidas em laboratórios,[73] foram levantadas dúvidas sobre se a síntese natural não biológica de RNA é possível.[74] As primeiras "ribozimas" podem ter sido formadas por ácidos nucleicos mais simples, como PNA, TNA ou GNA, que teriam sido substituídos posteriormente por RNA.[75][76]
Em 2003, foi proposto que precipitados de sulfeto de metal poroso ajudariam a síntese de RNA em cerca de 100 °C (212 °F) e pressões do fundo do oceano perto de fontes hidrotermais. Nessa hipótese, as membranas lipídicas seriam os últimos grandes componentes celulares a aparecer e até então, as proto-células estariam confinadas aos poros.[77]
Primeiro metabolismo: mundo ferro–enxofre
editarUma série de experimentos iniciados em 1997 mostrou que os estágios iniciais na formação de proteínas a partir de materiais inorgânicos, incluindo monóxido de carbono e sulfeto de hidrogênio, poderiam ser alcançados usando sulfeto de ferro e sulfeto de níquel como catalisadores. A maioria das etapas exigia temperaturas de cerca de 100 °C (212 °F) e pressões moderadas, embora um estágio exigisse 250 °C (482 °F) e uma pressão equivalente à encontrada abaixo de 7 km (4.3 mi) de rocha. Portanto, foi sugerido que a síntese autossustentável de proteínas poderia ter ocorrido perto de fontes hidrotermais.[78]
Primeiras membranas: mundo lipídico
editar
Foi sugerido que "bolhas" de parede dupla de lipídios, como aquelas que formam as membranas externas das células, podem ter sido uma primeira etapa essencial.[79] Experimentos que simularam as condições da Terra primitiva relataram a formação de lipídios, e estes podem formar lipossomas espontaneamente, "bolhas" de parede dupla, e então se reproduzir. Embora eles não sejam intrinsecamente portadores de informações como os ácidos nucleicos, eles estariam sujeitos à seleção natural para longevidade e reprodução. Os ácidos nucleicos, como o RNA, podem então ter se formado mais facilmente dentro dos lipossomas do que fora deles.[80]
Hipótese da argila
editarO RNA é complexo e há dúvidas se ele pode ser produzido de forma não biológica na natureza.[74] Algumas argilas, notadamente a montmorillonita, têm propriedades que as tornam aceleradores plausíveis para o surgimento de um mundo de RNA: elas crescem por autor-replicação de seu padrão cristalino; eles estão sujeitos a um análogo da seleção natural, à medida que a "espécie" de argila que cresce mais rápido em um ambiente particular rapidamente se torna dominante, e podem catalisar a formação de moléculas de RNA.[81] Embora essa ideia não tenha se tornado o consenso científico, ela ainda tem defensores ativos.[82]
Pesquisa em 2003 relatou que a montmorilonita também pode acelerar a conversão de ácidos graxos em "bolhas" e que as "bolhas" podem encapsular o RNA anexado à argila. Essas "bolhas" podem então crescer absorvendo lipídios adicionais e então se dividir. A formação das primeiras células pode ter sido auxiliada por processos semelhantes.[83]
Uma hipótese semelhante apresenta argilas ricas em ferro autorreplicantes como as progenitoras de nucleotídeos, lipídios e aminoácidos.[84]
Impacto ambiental e evolutiva dos tapetes microbianos
editar
Esteiras microbianas são colônias multicamadas e multiespécies de bactérias e outros organismos que geralmente têm apenas alguns milímetros de espessura, mas ainda contêm uma ampla variedade de ambientes químicos, cada um favorecendo um conjunto diferente de microrganismos.[85] Até certo ponto, cada esteira forma sua própria cadeia alimentar, já que os subprodutos de cada grupo de microrganismos geralmente servem como "alimento" para grupos adjacentes.[86]
Estromatólitos são pilares atarracados construídos como microorganismos em esteiras que migram lentamente para cima para evitar serem sufocados por sedimentos depositados neles pela água. Houve um debate vigoroso sobre a validade de alegados fósseis anteriores a 3 Ga,[87] com os críticos argumentando que os chamados estromatólitos poderiam ter sido formados por processos não biológicos. Em 2006, outro achado de estromatólitos foi relatado na mesma parte da Austrália que os anteriores, em rochas datadas de 3,5 Ga.[88]
Em tapetes subaquáticos modernos, a camada superior geralmente consiste em cianobactérias fotossintetizantes que criam um ambiente rico em oxigênio, enquanto a camada inferior é livre de oxigênio e frequentemente dominada pelo sulfeto de hidrogênio emitido pelos organismos que vivem ali.[86] Estima-se que o aparecimento de fotossíntese oxigenada por bactérias em tapetes aumentou a produtividade biológica por um fator entre 100 e 1 000. O agente redutor usado pela fotossíntese oxigenada é a água, que é muito mais abundante do que os agentes redutores produzidos geologicamente exigidos pela fotossíntese não oxigenada anterior.[89] Desse ponto em diante, a própria vida produziu significativamente mais dos recursos de que precisava do que os processos geoquímicos.[90] O oxigênio é tóxico para organismos que não estão adaptados a ele, mas aumenta muito a eficiência metabólica dos organismos adaptados ao oxigênio.[91][92] O oxigênio se tornou um componente significativo da atmosfera da Terra em cerca de 2,4 Ga.[93] Embora os eucariotos possam ter existido muito antes,[94][95] a oxigenação da atmosfera foi um pré-requisito para a evolução da maioria células eucarióticas complexas, a partir das quais todos os organismos multicelulares são construídos. A fronteira entre as camadas ricas em oxigênio e as livres de oxigênio nas esteiras microbianas teria se movido para cima quando a fotossíntese foi encerrada durante a noite, e então para baixo quando foi retomada no dia seguinte.[96] Isso teria criado uma pressão de seleção para que os organismos desta zona intermediária adquirissem a capacidade de tolerar e usar o oxigênio, possivelmente via endossimbiose, onde um organismo vive dentro de outro e ambos se beneficiam de sua associação.[19]
As cianobactérias possuem os "kits de ferramentas" bioquímicos mais completos de todos os organismos formadores de esteira. Conseqüentemente, eles são os mais autossuficientes dos organismos de esteira e foram bem adaptados para atacar por conta própria tanto como esteiras flutuantes quanto como o primeiro fitoplâncton, fornecendo a base da maioria das cadeias alimentares marinhas.[19]
Diversificação dos eucariontes
editar| Eukarya |
| |||||||||||||||||||||||||||||||||
Cromatina, núcleo, sistema endomembranoso e mitocôndrias
editarOs eucariotos podem estar presentes muito antes da oxigenação da atmosfera,[94] mas a maioria dos eucariotos modernos requer oxigênio, que suas mitocôndrias usam para alimentar a produção de ATP, o suprimento interno de energia de todas as células conhecidas.[96] Na década de 1970, foi proposto e após muito debate, amplamente aceito que os eucariotos surgiram como resultado de uma sequência de endossimbiose entre procariotos. Por exemplo: um micro-organismo predatório invadiu um grande procarionte, provavelmente um arqueano, mas o ataque foi neutralizado, e o atacante fixou residência e evoluiu para a primeira das mitocôndrias; uma dessas quimeras tentou mais tarde engolir uma cianobactéria fotossintetizante, mas a vítima sobreviveu dentro do atacante e a nova combinação se tornou a ancestral das plantas; e assim por diante. Após o início de cada endossimbiose, os parceiros teriam eliminado a duplicação improdutiva das funções genéticas reorganizando seus genomas, um processo que às vezes envolvia a transferência de genes entre eles.[99][100][101] Outra hipótese propõe que as mitocôndrias eram originalmente endossimbiontes que metabolizam enxofre ou hidrogênio e se tornaram consumidoras de oxigênio posteriormente.[102] Por outro lado, as mitocôndrias podem ter feito parte do equipamento original dos eucariotos.[103]
Há um debate sobre quando os eucariotos apareceram pela primeira vez: a presença de esteranos em folhelhos australianos pode indicar que os eucariotos estavam presentes 2,7 Ga;[95] no entanto, uma análise em 2008 concluiu que esses produtos químicos se infiltraram nas rochas com menos de 2,2 Ga e não provam nada sobre as origens dos eucariotos.[104] Fósseis da alga Grypania foram relatados em rochas de 1,85 bilhão de anos (originalmente datadas de 2,1 Ga, mas posteriormente revisadas[105]), e indica que os eucariotos com organelas já haviam evoluído.[106] Uma coleção diversa de fósseis de algas foi encontrada em rochas datadas entre 1,5 e 1,4 Ga.[107] Os primeiros fósseis de fungos conhecidos datam de 1,43 Ga.[108]
Plastídios
editarPensa-se que os plastídios, a superclasse de organelas das quais os cloroplastos são os exemplares mais conhecidos, tenham se originado de cianobactérias endossimbióticas. A simbiose evoluiu em torno de 1,5 Ga e permitiu que os eucariotos realizassem a fotossíntese oxigenada.[96] Três linhagens evolutivas de plastídios fotossintéticos surgiram desde então, nas quais os plastídios são nomeados de forma diferente: cloroplastos em algas verdes e plantas, rodoplastos em algas vermelhas e cianelas nas glaucófitas.[109]
Reprodução sexual e organismos multicelulares
editarEvolução da reprodução sexual
editarAs características definidoras da reprodução sexuada em eucariotos são a meiose e a fertilização. Nesse tipo de reprodução há muita recombinação genética, na qual os descendentes recebem 50% dos genes de cada um dos pais,[110] ao contrário da reprodução assexuada, na qual não há recombinação. As bactérias também trocam DNA por conjugação bacteriana, com a qual podem adquirir, por exemplo, genes de resistência a antibióticos e outras toxinas e a capacidade de usar novos metabólitos.[111] No entanto, a conjugação não é um meio de reprodução e não se limita a membros da mesma espécie. Existem casos em que bactérias transferem DNA para plantas e animais.[112]
Por outro lado, a transformação bacteriana é claramente uma adaptação para a transferência de DNA entre bactérias da mesma espécie. A transformação bacteriana é um processo complexo que envolve os produtos de vários genes bacterianos e pode ser considerada uma forma de sexo em bactérias.[113][114] Este processo ocorre naturalmente em pelo menos 67 espécies procarióticas (de sete filos diferentes).[115] A reprodução sexual em eucariotos pode ter evoluído a partir da transformação bacteriana.[116]
As desvantagens da reprodução sexual são bem conhecidas: a redistribuição genética da recombinação pode resultar em combinações de genes favoráveis; e como os machos não aumentam diretamente o número de descendentes na próxima geração, uma população assexuada pode ultrapassar o número de descendentes e deslocar em apenas 50 gerações uma população sexual que é igual em todos os outros aspectos.[110] No entanto, a grande maioria dos animais, plantas, fungos e protistas reproduzem-se sexualmente. Há fortes evidências de que a reprodução sexual se originou muito cedo na história dos eucariotos e que os genes que a controlam mudaram muito pouco desde então.[117] Explicar como a reprodução sexual evoluiu e sobreviveu é um quebra-cabeça não resolvido.[118]

A Hipótese da Rainha Vermelha indica que a reprodução sexuada fornece proteção contra parasitas, porque é mais fácil para os parasitas evoluírem meios para superar as defesas de clones geneticamente idênticos do que de espécies sexuais com alvos móveis, e há algumas evidências experimentais para isso. No entanto, ainda há dúvidas se isso explicaria a sobrevivência das espécies sexuais se várias espécies de clones semelhantes estivessem presentes, já que um dos clones pode sobreviver a ataques de parasitas por tempo suficiente para superar as espécies sexuais.[110] Além disso, ao contrário do que é esperado pela hipótese da Rainha Vermelha, Kathryn A. Hanley et al. eles descobriram que a prevalência, abundância e intensidade média de parasitas de ácaros foram significativamente maiores em controles sexuais do que em assexuais que vivem no mesmo habitat.[120] Além disso, o biólogo Matthew Parker, após revisar muitos estudos genéticos sobre resistência a doenças de plantas, não conseguiu encontrar um único exemplo consistente com o conceito de que os patógenos são o principal agente seletivo responsável pela reprodução sexual no hospedeiro.[121]
A hipótese da mutação determinística de Alexey Kondrashov sustenta que cada organismo tem mais de uma mutação prejudicial, e os efeitos combinados dessas mutações são mais prejudiciais do que a soma dos danos causados por cada mutação separadamente. Nesse caso, a recombinação sexual de genes reduzirá o dano que as mutações ruins causam aos descendentes, e ao mesmo tempo, removerá algumas mutações ruins do fundo genético, isolando-as em indivíduos que morrem rapidamente porque têm um número acima da média de mutações ruins. No entanto, as evidências sugerem que os postulados da hipótese de mutação determinística são fracos, porque muitas espécies têm em média menos de uma mutação prejudicial por indivíduo e nenhuma espécie que foi investigada mostra evidências de sinergia entre mutações prejudiciais.[110]
A natureza aleatória da recombinação faz com que a abundância relativa de características alternativas varie de geração para geração. Essa deriva genética sozinha é insuficiente para tornar a reprodução sexual vantajosa, mas uma combinação de deriva genética e seleção natural pode ser suficiente. Quando o acaso produz combinações de boas características, a seleção natural oferece uma grande vantagem às linhagens nas quais essas características estão geneticamente ligadas. Por outro lado, os benefícios das boas características são neutralizados se aparecerem junto com as más. A recombinação sexual dá boas características às oportunidades de serem vinculadas a outras boas características, e os modelos matemáticos sugerem que isso pode ser mais do que suficiente para compensar as desvantagens da reprodução sexual.[118] Outras combinações de hipóteses que são inadequadas separadamente também foram examinadas.[110]
A função adaptativa do sexo continua sendo uma questão importante não resolvida hoje. Modelos concorrentes para explicar a função adaptativa do sexo foram revisados por John A. Birdsell e Christopher Wills.[122] Todas as hipóteses discutidas acima dependem dos possíveis efeitos benéficos da variação genética aleatória produzida pela recombinação genética. Uma concepção alternativa é que o sexo se originou, e tem sido mantido, como um processo de reparo do DNA danificado, e que a variação genética produzida é um subproduto ocasionalmente benéfico.[116][123]
Multicelularidade
editarDefinições mais simples de "multicelular" ou "multicelular", por exemplo "tendo células múltiplas", podem incluir cianobactérias coloniais, como Nostoc. Mesmo uma definição técnica como "ter o mesmo genoma, mas diferentes tipos de células" ainda incluiria alguns gêneros de algas verdes Volvox, que têm células especializadas em reprodução.[124] A multicelularidade evoluiu independentemente em organismos tão diversos quanto esponjas e outros animais, fungos, plantas, algas marrons, cianobactérias, fungos mucilaginosos e mixobactérias.[105][125] Para não tornar este artigo muito extenso, este artigo concentra-se em organismos que apresentam a maior especialização de células e variedade de tipos de células, embora esta abordagem da evolução da complexidade biológica possa ser considerada "bastante antropocêntrica".[126]

A vantagem inicial da multicelularidade pode ser devido a: compartilhamento mais eficiente de nutrientes que são digeridos fora da célula,[128] aumentando a resistência a predadores, muitos dos quais atacados por fagocitização; a capacidade de suportar correntes ao aderir a uma superfície firme; a capacidade de chegar mais alto para alimentar por filtração ou obter luz para a fotossíntese;[129] a capacidade de criar um ambiente interno que forneça proteção contra o ambiente externo;[126] e até mesmo a oportunidade de formar um grupo de células. "de forma inteligente", compartilhando informações.[127] Essas características também forneceriam oportunidades para outros organismos se diversificarem, criando ambientes mais diversos do que as esteiras microbianas poderiam gerar.[129]
A multicelularidade com células diferenciadas é benéfica para o organismo como um todo, mas desvantajosa do ponto de vista das células individuais, muitas das quais perderam a oportunidade de se reproduzir por conta própria. Em um organismo multicelular assexuado, células indisciplinadas que mantêm a capacidade de reprodução podem assumir o controle e reduzir o organismo a uma massa celular indiferenciada. A reprodução sexual elimina essas células indisciplinadas da próxima geração e, portanto, parece ser um requisito para a multicelularidade complexa.[129]
As evidências disponíveis indicam que os eucariotos evoluíram muito antes, mas permaneceram despercebidos até a rápida diversificação, por volta de 1 000 milhões de anos atrás. A única coisa em que os eucariotos claramente superam o número de bactérias e arquéias é sua capacidade de assumir várias formas, e a reprodução sexual permitiu que os eucariotos tirassem proveito disso para produzir organismos com células múltiplas que diferem em forma e função.[129]
Ao comparar a composição de famílias de fatores de transcrição e motivos de rede regulatória entre organismos unicelulares e organismos multicelulares, os cientistas descobriram que existem muitas novas famílias de fatores de transcrição e três novos tipos de motivos de rede regulatória em organismos multicelulares, e as novas famílias de transcrição fatores são preferencialmente conectados nesses novos motivos de rede, que são essenciais para o desenvolvimento multicelular. Esses resultados propõem um mecanismo plausível para a contribuição de novas famílias de fatores de transcrição e novos motivos de rede para a origem de organismos multicelulares no nível regulatório da transcrição.[130]
Evidência fóssil
editarOs fósseis da biota franceviliana, datados de 2 100 Ma, são os organismos fósseis mais antigos conhecidos que são claramente multicelulares.[35] Eles podem ter células diferenciadas.[131] Outro fóssil multicelular muito antigo é Qingshania, datado de 1 700 Ma, que parece consistir em células virtualmente idênticas. A alga vermelha chamada Bangiomorpha, datada de 1 200 Ma, é o primeiro organismo conhecido a ter células diferenciadas especializadas com certeza, e também é o organismo mais antigo conhecido a se reproduzir sexualmente.[129] Fósseis de 1 430 Ma interpretados como fungos parecem ter sido multicelulares com células diferenciadas.[108] O organismo "cadeia de dois" Horodyskia, encontrado em rochas que datam de 1 500 Ma a 900 Ma, pode ter sido um dos primeiros metazoários,[105] no entanto, também foi interpretado como um foraminífero colonial.[119]
Emergência de animais
editar
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
Os animais são eucariotos multicelulares,[nota 1] e se distinguem das plantas, algas e fungos por não possuírem paredes celulares.[134] Todos os animais são móveis,[135] mesmo que apenas em certos estágios da vida. Todos os animais, exceto as esponjas, têm corpos diferenciados em tecidos separados, incluindo músculos, que movem partes do animal ao se contrair, e tecido nervoso, que transmite e processa sinais.[136] Em novembro de 2019, pesquisadores relataram a descoberta da Caveasphaera, um organismo multicelular encontrado em rochas de 609 milhões de anos, que não é facilmente definido como um animal ou não animal, o que pode estar relacionado a uma das primeiras instâncias de animal evolução.[137][138] Estudos de fósseis de Caveasphaera sugeriram que o desenvolvimento embrionário semelhante ao animal surgiu muito antes dos fósseis de animais claramente definidos mais antigos.[137] e pode ser consistente com estudos que sugerem que a evolução animal pode ter começado há cerca de 750 milhões de anos.[138][139]
No entanto, os primeiros fósseis de animais amplamente aceitos são os cnidários de aparência bastante moderna (o grupo que inclui águas-vivas, anêmonas do mar e Hydra), possivelmente de cerca de 580 Ma, embora os fósseis da Formação Doushantuo só possam ser datados aproximadamente. Sua presença implica que as linhagens cnidária e bilateriana já haviam divergido.[140]
A biota Ediacara, que floresceu nos últimos 40 milhões de anos antes do início do Cambriano,[141] foram os primeiros animais com mais de alguns centímetros de comprimento. Muitos eram planos e tinham uma aparência "acolchoada" e pareciam tão estranhos que houve uma proposta para classificá-los como um reino separado, Vendozoa.[142] Outros, no entanto, foram interpretados como moluscos primitivos (Kimberella[143][144]), equinodermos (Arkarua[145]) e artrópodes (Spriggina,[146] Parvancorina[147]). Ainda há debate sobre a classificação desses espécimes, principalmente porque as características diagnósticas que permitem aos taxonomistas classificar organismos mais recentes, como semelhanças com organismos vivos, geralmente estão ausentes nos Ediacarans. No entanto, parece haver pouca dúvida de que Kimberella' era pelo menos um animal bilateriano triploblástico, em outras palavras, um animal significativamente mais complexo do que os cnidários.[148]
A pequena fauna de conchas é uma coleção muito mista de fósseis encontrados entre os períodos Ediacariano Superior e Cambriano Médio. O mais antigo, Cloudina, mostra sinais de defesa bem-sucedida contra a predação e pode indicar o início de uma corrida armamentista evolutiva. Algumas minúsculas conchas do Cambriano primitivo quase certamente pertenciam a moluscos, enquanto os proprietários de algumas "placas de armadura", Halkieria e Microdictyon, foram eventualmente identificados quando espécimes mais completos foram encontrados em lagerstätte cambriano que preservava animais de corpo mole.[149]

Na década de 1970, já havia um debate sobre se o surgimento dos filos modernos era "explosivo" ou gradual, mas oculto pela escassez de fósseis de animais pré-cambrianos.[149] Uma reanálise de fósseis do lagerstätte Folhelho Burgess aumentou o interesse no assunto ao revelar animais, como Opabinia, que não se encaixavam em nenhum filo conhecido. Na época, isso foi interpretado como evidência de que os filos modernos evoluíram muito rapidamente na explosão cambriana e que as "maravilhas estranhas" do xisto de Burgess mostraram que o Cambriano Inferior foi um período exclusivamente experimental de evolução animal.[151] Descobertas posteriores de animais semelhantes e o desenvolvimento de novas abordagens teóricas levaram à conclusão de que muitas das "maravilhas estranhas" eram "tias" ou "primas" evolucionárias de grupos modernos[152]—por exemplo, que Opabinia era um membro dos lobópodes, um grupo que inclui os ancestrais dos artrópodes, e que pode ter sido intimamente relacionado aos tardígrados modernos.[153] No entanto, ainda há muito debate sobre se a explosão cambriana foi realmente explosiva e, em caso afirmativo, como e por que aconteceu e por que parece única na história dos animais.[154]
Deuterostômios e os primeiros vertebrados
editar
A maioria dos animais no centro do debate sobre a explosão cambriana são [[Protostomia|protostômios], um dos dois grupos principais de animais complexos. O outro grupo principal, os deuterostômios, contém invertebrados como estrelas do mar e ouriços-do-mar (equinodermos), bem como cordados (veja abaixo). Muitos equinodermos têm "conchas" duras de calcita, que são bastante comuns da pequena fauna de conchas do início do Cambriano em diante.[149] Outros grupos de deuterostômios são de corpo mole, e a maioria dos fósseis de deuterostômios cambrianos significativos vêm da fauna de Chengjiang, um lagerstätte na China.[156] Os cordados são outro grupo importante de deuterostômios: animais com um cordão nervoso dorsal distinto. Os cordados incluem invertebrados de corpo mole, como tunicados, bem como vertebrados—animais com espinha dorsal. Enquanto os fósseis de tunicado são anteriores à explosão cambriana,[157] os fósseis de Chengjiang Haikouichthys e Myllokunmingia parecem ser verdadeiros vertebrados,[25] e Haikouichthys tinha vértebras distintas, que podem ter sido ligeiramente mineralizadas.[158] Vertebrados com mandíbulas, como os acantodianos, apareceram pela primeira vez no Ordoviciano Superior.[159]
Colonização da terra
editarA adaptação à vida na terra é um grande desafio: todos os organismos terrestres precisam evitar o ressecamento e todos aqueles acima do tamanho microscópico devem criar estruturas especiais para suportar a gravidade; os sistemas de respiração e troca gasosa precisam mudar; sistemas reprodutivos não podem depender de água para transportar ovos e espermas uns para os outros.[160][161][162] Embora as primeiras evidências de plantas e animais terrestres datem do período Ordoviciano (488 a 444 Ma), e uma série de linhagens de micro-organismos tenham chegado à terra muito antes,[163][164] os ecossistemas terrestres modernos só apareceram no final do Devoniano, cerca de 385 a 359 Ma.[165] Em maio de 2017, a evidência da vida mais antiga conhecida na terra pode ter sido encontrada em géiserita de 3,48 bilhões de anos e outros depósitos minerais relacionados (frequentemente encontrados em torno de fontes termais e gêiseres) descobertos no Cráton de Pilbara, na Austrália Ocidental.[166][167] Em julho de 2018, os cientistas relataram que a vida mais antiga na terra pode ter sido bactérias que viviam na terra há 3,22 bilhões de anos.[168] Em maio de 2019, cientistas relataram a descoberta de um fungo fossilizado, chamado Ourasphaira giraldae, no Ártico canadense, que pode ter crescido em terras há um bilhão de anos, bem antes das plantas viverem na terra.[169][170][171]
Evolução de antioxidantes terrestres
editarO oxigênio é um potente oxidante cujo acúmulo na atmosfera terrestre resultou do desenvolvimento da fotossíntese ao longo de 3 Ga, em cianobactérias (algas verde-azuladas), que foram os organismos fotossintéticos oxigenados mais primitivos. As algas marrons acumulam antioxidantes minerais inorgânicos como rubídio, vanádio, zinco, ferro, cobre, molibdênio, selênio e iodo, que se concentra mais de 30 000 vezes a concentração deste elemento na água do mar. Enzimas antioxidantes endógenas protetoras e antioxidantes dietéticos exógenos ajudaram a prevenir o dano oxidativo. A maioria dos antioxidantes minerais marinhos atua nas células como oligoelementos essenciais em oxirredução de metaloenzimas e antioxidantes.
Quando as plantas e animais começaram a entrar nos rios e pousar por volta de 500 Ma, a deficiência ambiental desses antioxidantes minerais marinhos foi um desafio para a evolução da vida terrestre.[172][173] As plantas terrestres otimizaram lentamente a produção de “novos” antioxidantes endógenos, como ácido ascórbico, polifenol, Flavonoide, tocoferol, etc. Alguns destes apareceram mais recentemente, nos últimos 200–50 Ma, em frutas se flores de plantas angiospérmicas.
Na verdade, as angiospermas (o tipo dominante de planta hoje) e a maioria de seus pigmentos antioxidantes evoluíram durante o período Jurássico Superior. As plantas empregam antioxidantes para defender suas estruturas contra espécies reativas de oxigênio produzidas durante a fotossíntese. Os animais estão expostos aos mesmos oxidantes e desenvolveram sistemas antioxidantes enzimáticos endógenos.[174] Iodo na forma de íon iodeto I é o elemento essencial rico em elétrons mais primitivo e abundante na dieta de organismos marinhos e terrestres, e o iodeto atua como um doador de elétrons e possui este antioxidante ancestral funcionam em todas as células concentradoras de iodeto, desde algas marinhas primitivas até vertebrados terrestres mais recentes.[175]
Evolução do solo
editarAntes da colonização da terra, o solo, uma combinação de partículas minerais e matéria orgânica decomposta, não existia. As superfícies da terra seriam de rocha nua ou areia instável produzida pelo intemperismo. A água e quaisquer nutrientes nela teriam sido drenados muito rapidamente.[165] Na peneplanície sub-câmbrica na Suécia, por exemplo, a profundidade máxima de caulinitização por intemperismo no Neoproterozoico é de cerca de 5 m, em contraste, os depósitos de caulim próximos desenvolvidos no Mesozoico são muito mais espessos.[176] Argumentou-se que no final do Neoproterozoico a lavagem de lençóis era um processo dominante de erosão do material de superfície devido à falta de plantas na terra.[177]

Biofilme de cianobactérias, que não são plantas, mas usam os mesmos mecanismos de fotossíntese, foram encontrados em desertos modernos e apenas em áreas inadequadas para plantas vasculares. Isso sugere que esteiras microbianas podem ter sido os primeiros organismos a colonizar a terra seca, possivelmente no Pré-Cambriano. As cianobactérias formadoras de esteira podem ter gradualmente desenvolvido resistência à dessecação à medida que se espalham dos mares para as zonas costeiras e depois para a terra.[165] Os líquens, que são combinações simbióticas de um fungo (quase sempre um ascomiceto) e um ou mais fotossintetizadores (algas verdes ou cianobactérias),[178] também são colonizadores importantes de ambientes sem vida,[165] e sua capacidade de quebrar rochas contribui para formação do solo em situações em que as plantas não sobrevivem.[178] Os primeiros fósseis de ascomicetos conhecidos datam de 423 a 419 Ma no Siluriano.[165]
A formação do solo teria sido muito lenta até o aparecimento de animais escavadores, que misturam os componentes minerais e orgânicos do solo e cujas fezes são a principal fonte dos componentes orgânicos.[165] Tocas foram encontradas em sedimentos ordovicianos e são atribuídas a anelídeos ("vermes") ou artrópodes.[165][179]
Plantas e a crise de madeira no Devoniano Médio
editar

Nas algas aquáticas, quase todas as células são capazes de fotossíntese e são quase independentes. A vida na terra exigia que as plantas se tornassem internamente mais complexas e especializadas: a fotossíntese era mais eficiente no topo; raízes eram necessárias para extrair água do solo; as partes intermediárias tornaram-se suportes e sistemas de transporte de água e nutrientes.[160][180]
Esporos de plantas terrestres, possivelmente como hepáticas, foram encontrados em rochas do Ordoviciano Médio datadas de cerca de 476 milhões de anos. Nas rochas do Siluriano Médio de 430 Ma, existem fósseis de plantas reais, incluindo musgos como Baragwanathia; a maioria tinha menos de 10 cm (3.9 in) e alguns parecem intimamente relacionados às plantas vasculares, o grupo que inclui as árvores.[180]
No final do Devoniano 370 Ma, árvores como Archaeopteris eram tão abundantes que mudaram os sistemas fluviais de quase todos entrelaçados para quase sempre sinuosos, porque suas raízes prendiam o solo com firmeza.[181] Na verdade, eles causaram a "crise da madeira do Devoniano Tardio"[182] porque:
- Eles removeram mais dióxido de carbono da atmosfera, reduzindo o efeito estufa e causando uma era do gelo no período Carbonífero.[183] Em ecossistemas posteriores, o dióxido de carbono "preso" na madeira é devolvido à atmosfera pela decomposição da madeira morta. No entanto, a evidência fóssil mais antiga de fungos que podem decompor a madeira também vem do Devoniano Tardio.[184]
- O aumento da profundidade das raízes das plantas levou a uma maior lavagem de nutrientes nos rios e mares pela chuva. Isso causou o florescimento de algas cujo alto consumo de oxigênio causou eventos anóxicos em águas mais profundas, aumentando a taxa de extinção entre os animais de águas profundas.[183]
Invertebrados terrestres
editarOs animais tiveram que mudar seus sistemas de alimentação e excretor, e a maioria dos animais terrestres desenvolveu a fertilização interna de seus ovos.[162] A diferença no índice de refração entre a água e o ar exigia mudanças em seus olhos. Por outro lado, de certa forma, o movimento e a respiração tornaram-se mais fáceis, e a melhor transmissão de sons de alta frequência no ar estimulou o desenvolvimento da audição.[161]

O mais antigo animal que respira ar é o Pneumodesmus, um arquipolipodano milípede do Siluriano Médio, com cerca de 428 milhões de anos.[185][186] Sua natureza terrestre com respiração aérea é evidenciada pela presença de espiráculos, as aberturas dos sistemas traqueais.[187] No entanto, alguns vestígios de fósseis anteriores da fronteira Cambriana-Ordoviciana cerca de 490 Ma são interpretados como rastros de grandes artrópodes anfíbios em dunas de areia costeiras e podem ter sido feitos por euticarcinoides,[188] que são consideradas "tias" evolucionárias de miriápodes.[189] Outros vestígios de fósseis do Ordoviciano Superior com pouco mais de 445 Ma provavelmente representam invertebrados terrestres, e há evidências claras de numerosos artrópodes nas costas e planícies aluviais pouco antes da fronteira Siluriana-Devoniana, cerca de 415 Ma, incluindo sinais de que alguns artrópodes comeu plantas.[190] Os artrópodes foram bem pré-adaptados para colonizar a terra, porque seus exoesqueletos articulados existentes forneciam proteção contra a dessecação, suporte contra a gravidade e um meio de locomoção que não dependia de água.[162][191]
O registro fóssil de outros grupos importantes de invertebrados em terra é pobre: nenhum para vermes planas não parasitas, nematóides ou nemertinos; alguns nematóides parasitas foram fossilizados em âmbar; fósseis de vermes anelídeos são conhecidos do Carbonífero, mas eles ainda podem ter sido animais aquáticos; os fósseis mais antigos de gastrópodes em terra datam do Carbonífero Superior, e esse grupo pode ter tido que esperar até que a camada de folhas se tornasse abundante o suficiente para fornecer as condições de umidade de que precisam.[161]
Os primeiros fósseis confirmados de insetos voadores datam do Carbonífero Superior, mas acredita-se que os insetos desenvolveram a capacidade de voar no Carbonífero Inferior ou mesmo no Devoniano Superior. Isso lhes deu uma ampla gama de nichos ecológicos para alimentação e reprodução, e um meio de escapar de predadores e de mudanças desfavoráveis no ambiente.[192] Cerca de 99% das espécies de insetos modernos voam ou são descendentes de espécies voadoras.[193]
Primeiros vertebrados terrestres
editar
Os tetrápodes, vertebrados com quatro membros, evoluíram de outros peixes rhipidistianos em um período de tempo relativamente curto durante o Devoniano Superior (370 a 360 Ma).[196] Os primeiros grupos são agrupados como labirintodonte. Eles mantiveram girinos aquáticos semelhantes a alevins, um sistema ainda visto em anfíbios modernos.
O iodo e o T4/T3 estimulam a metamorfose anfíbia e a evolução do sistema nervoso transformando o girino aquático vegetariano em sapo carnívoro "mais evoluído" terrestre com melhores habilidades neurológicas, visuoespaciais, olfativas e cognitivas para a caça.[172] A nova ação hormonal do T3 foi possibilitada pela formação de receptores T3 nas células dos vertebrados. Primeiramente, cerca de 600-500 milhões de anos atrás, na primitiva Chordata apareceram os receptores alfa T3 com ação metamorfosante e então, cerca de 250-150 milhões de anos atrás, nas aves e Mammalia apareceram os receptores beta T3 com ações metabólicas e termogenéticas.[197]
Dos anos 1950 ao início dos anos 1980, pensava-se que os tetrápodes evoluíram de peixes que já haviam adquirido a capacidade de rastejar na terra, possivelmente para ir de um lago que estava secando para outro mais profundo. No entanto, em 1987, fósseis quase completos de Acanthostega de cerca de 363 Ma mostraram que este animal de transição do Devoniano Superior tinha pernas e pulmões e guelras, mas nunca poderia ter sobrevivido em terra: seus membros e suas articulações de pulso e tornozelo eram muito fracos para suportar seu peso; suas costelas eram curtas demais para evitar que seus pulmões fossem espremidos por seu peso; sua barbatana caudal parecida com a de um peixe teria sido danificada ao ser arrastada pelo solo. A hipótese atual é que o Acanthostega, que tinha cerca de 1 metro (3,3 pés) de comprimento, era um predador totalmente aquático que caçava em águas rasas. Seu esqueleto era diferente do da maioria dos peixes, de maneiras que lhe permitiam erguer a cabeça para respirar ar enquanto o corpo permanecia submerso, incluindo: suas mandíbulas mostram modificações que o teriam permitido engolir ar; os ossos na parte de trás do crânio estão travados, fornecendo fortes pontos de fixação para os músculos que levantaram sua cabeça; a cabeça não está ligada à cintura escapular e tem um pescoço distinto.[194]
A proliferação devoniana de plantas terrestres pode ajudar a explicar por que respirar ar teria sido uma vantagem: folhas caindo em riachos e rios teriam encorajado o crescimento da vegetação aquática; isso teria atraído invertebrados pastando e pequenos peixes que os atacavam; teriam sido presas atraentes, mas o ambiente não era adequado para os grandes peixes marinhos predadores; a respiração do ar teria sido necessária porque essas águas teriam carência de oxigênio, uma vez que a água quente retém menos oxigênio dissolvido do que a água marinha mais fria e uma vez que a decomposição da vegetação teria usado parte do oxigênio.[194]
Descobertas posteriores revelaram formas de transição anteriores entre o Acanthostega e animais totalmente semelhantes a peixes.[198] Infelizmente, há uma brecha (brecha de Romer) de cerca de 30 Ma entre os fósseis de tetrápodes ancestrais e fósseis de vertebrados do Carbonífero Médio que parecem bem adaptados para a vida na terra. Alguns deles se parecem com os primeiros parentes dos anfíbios modernos, a maioria dos quais precisam manter suas peles úmidas e colocar seus ovos na água, enquanto outros são aceitos como os primeiros parentes dos amniotas, cuja pele à prova d'água e membranas de ovo lhes permitem viver e raça longe da água.[195]
Dinossauros, aves e mamíferos
editar| Amniotas |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Os amniotas, cujos ovos podem sobreviver em ambientes secos, provavelmente evoluíram no período do Carbonífero Superior (330 a 298,9 Ma). Os primeiros fósseis dos dois grupos sobreviventes de amniota, sinapsídeos e sauropsídeos, datam de cerca de 313 milhões de anos.[200][201] Os pelicossauros sinapsídeos e seus descendentes, os terapsídeos, são os vertebrados terrestres mais comuns nos leitos fósseis mais conhecidos do Permiano (298,9 a 251,902 Ma). No entanto, na época, todas estavam em zonas temperadas em latitudes médias, e há evidências de que ambientes mais quentes e secos próximos ao Equador foram dominados por sauropsídeos e anfíbios. [202]
O evento de extinção Permiano-Triássico eliminou quase todos os vertebrados terrestres,[203] bem como a grande maioria de outras formas de vida.[204] Durante a lenta recuperação desta catástrofe, estimada em 30 milhões de anos,[205] um grupo sauropsídeo anteriormente obscuro tornou-se o mais abundante e diverso dos vertebrados terrestres: alguns fósseis de arquosauriformes ("formas de lagarto governantes") foram encontrados em rochas do Permiano Superior,[206] mas, no Triássico Médio, os arcossauros eram os vertebrados terrestres dominantes. Os dinossauros se distinguiram de outros arcossauros no final do Triássico e se tornaram os vertebrados terrestres dominantes dos períodos Jurássico e Cretáceo (201,3 a 66 Ma).[207]
Durante o Jurássico Superior, os pássaros evoluíram de pequenos dinossauros terópodes predadores.[208] Os primeiros pássaros herdaram dentes e longas caudas ósseas de seus ancestrais dinossauros,[208] mas alguns desenvolveram bicos córneos e desdentados no Jurássico Tardio[209] e caudas curtas de pigóstilo no Cretáceo Inferior.[210]
Enquanto os arcossauros e dinossauros estavam se tornando mais dominantes no Triássico, os sucessores em forma de mamífero dos terapsídeos evoluíram para pequenos insetívoros, principalmente noturnos. Este papel ecológico pode ter promovido a evolução dos mamíferos, por exemplo, a vida noturna pode ter acelerado o desenvolvimento da endotermia ("sangue quente") e cabelo ou pele.[211] Por volta de 195 Ma no Jurássico Inferior, havia animais que eram muito parecidos com os mamíferos de hoje em vários aspectos.[212] Infelizmente, há uma lacuna no registro fóssil em todo o Jurássico Médio.[213] No entanto, dentes fósseis descobertos em Madagáscar indicam que a divisão entre a linhagem que leva aos monotremados e a que leva a outros mamíferos vivos ocorreu por volta de 167 milhões de anos.[214] Depois de dominar nichos de vertebrados terrestres por cerca de 150 Ma, os dinossauros não aviários pereceram no evento de extinção Cretáceo-Paleógeno (66 Ma) junto com muitos outros grupos de organismos.[215] Os mamíferos ao longo do tempo dos dinossauros foram restritos a uma estreita faixa de taxa, tamanhos e formas, mas aumentaram rapidamente em tamanho e diversidade após a extinção,[216][217] com morcegos levando ao ar em 13 milhões de anos,[218] e os cetáceos ao mar em 15 milhões de anos.[219]
Plantas floridas
editar
|
|
As primeiras plantas com flores apareceram por volta de 130 Ma.[222] As 250 000 a 400 000 espécies de plantas com flores superam em número todas as outras plantas terrestres combinadas e são a vegetação dominante na maioria dos ecossistemas terrestres. Há evidências fósseis de que as plantas com flores se diversificaram rapidamente no Cretáceo Inferior, de 130 a 90 Ma,[220][221] e que sua ascensão foi associada à dos insetos polinizadores.[221] Entre as plantas com flores modernas, acredita-se que a Magnolia seja próxima do ancestral comum do grupo.[220] No entanto, os paleontólogos não conseguiram identificar os primeiros estágios na evolução das plantas com flores.[220][221]
Insetos sociais
editar
Os insetos sociais são notáveis porque a grande maioria dos indivíduos em cada colônia são estéreis. Isso parece contrário aos conceitos básicos de evolução, como a seleção natural e o gene egoísta. Na verdade, existem muito poucas espécies de insetos eussociais: apenas 15 das aproximadamente 2 600 famílias vivas de insetos contêm espécies eussociais, e parece que a eussocialidade evoluiu independentemente apenas 12 vezes entre os artrópodes, embora algumas linhagens eussociais tenham se diversificado em várias famílias. No entanto, os insetos sociais tiveram um sucesso espetacular; por exemplo, embora formigas e cupins representam apenas cerca de 2% das espécies de insetos conhecidas, eles formam mais de 50% da massa total dos insetos. Sua capacidade de controlar um território parece ser a base de seu sucesso.[223]
O sacrifício das oportunidades de procriação pela maioria dos indivíduos há muito tem sido explicado como consequência do método haplodiplóide incomum de determinação do sexo dessas espécies, que tem a consequência paradoxal de que duas filhas operárias estéreis da mesma rainha compartilham mais genes entre si do que com sua prole se eles pudessem procriar.[224] No entanto, EO Wilson e Bert Hölldobler argumentam que esta explicação é falha: por exemplo, é baseada na seleção de parentesco, mas não há evidência de nepotismo em colônias que possuem múltiplas rainhas. Em vez disso, eles escrevem, a eussocialidade evolui apenas em espécies que estão sob forte pressão de predadores e competidores, mas em ambientes onde é possível construir "fortalezas"; depois que as colônias estabeleceram essa segurança, elas ganham outras vantagens por meio do forrageamento cooperativo. Em apoio a essa explicação, eles citam o aparecimento de eussocialidade em ratos-toupeira batiérgidos,[223] que não são haplodiplóides.[225]
Os primeiros fósseis de insetos foram encontrados em rochas do Devoniano Inferior de cerca de 400 Ma, que preservam apenas algumas variedades de insetos que não voam. Os lagerstätten de Mazon Creek do Carbonífero Superior, cerca de 300 Ma, incluem cerca de 200 espécies, algumas gigantescas para os padrões modernos, e indicam que os insetos ocuparam seus principais nichos ecológicos modernos como herbívoros, detritívoros e insetívoros. Os cupins e formigas sociais aparecem pela primeira vez no Cretáceo Inferior, e abelhas sociais avançadas foram encontradas nas rochas do Cretáceo Superior, mas não se tornaram abundantes até o Cenozoico Médio.[226]
Humanos
editarA ideia de que, junto com outras formas de vida, os humanos modernos evoluíram de um ancestral comum antigo foi proposta por Robert Chambers em 1844 e adotada por Charles Darwin em 1871.[227] Os humanos modernos evoluíram de uma linhagem de andar ereto macacos que foram rastreados ao longo de 6 Ma até o Sahelanthropus.[228] As primeiras ferramentas de pedra conhecidas foram feitas cerca de 2,5 Ma, aparentemente por Australopithecus garhi, e foram encontradas perto de ossos de animais que apresentam arranhões feitos por essas ferramentas.[229] Os primeiros hominídeos tinhamcérebros do tamanho de chimpanzés, mas houve um aumento de quatro vezes nos últimos 3 milhões de anos; uma análise estatística sugere que o tamanho do cérebro hominídeo depende quase completamente da data dos fósseis, enquanto a espécie a que foram atribuídos tem apenas uma pequena influência.[230] Há um longo debate sobre se os humanos modernos evoluíram em todo o mundo simultaneamente a partir de hominíneos avançados existentes ou são descendentes de uma única pequena população na África, que então migrou para todo o mundo há menos de 200 000 anos e substituiu os anteriores espécies hominíneas.[231] Também há debate sobre se os humanos anatomicamente modernos tiveram um "Grande Salto para a Frente" intelectual, cultural e tecnológicomenos de 100 000 anos atrás, e em caso afirmativo, se isso foi devido a alterações neurológicas que não são visíveis nos fósseis.[232]
Extinções em massa
editar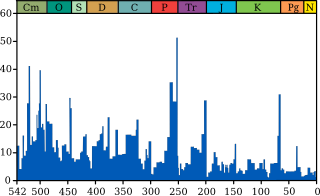
{kind=link}
A vida na Terra sofreu extinções em massa ocasionais pelo menos desde 542 milhões de anos. Embora fossem desastres na época, as extinções em massa às vezes aceleraram a evolução da vida na Terra. Quando o domínio de nichos ecológicos específicos passa de um grupo de organismos para outro, raramente é porque o novo grupo dominante é "superior" ao antigo e geralmente porque um evento de extinção elimina o antigo grupo dominante e abre caminho para o novo.[233]
O registro fóssil parece mostrar que as lacunas entre as extinções em massa estão se tornando mais longas e as taxas médias e de fundo de extinção estão diminuindo. Ambos os fenômenos podem ser explicados de uma ou mais maneiras:[234]
- Os oceanos podem ter se tornado mais hospitaleiros para a vida nos últimos 500 milhões de anos e menos vulneráveis a extinções em massa: o oxigênio dissolvido se espalhou e penetrou em maiores profundidades; o desenvolvimento da vida na terra reduziu o escoamento de nutrientes, e portanto, o risco de eutrofização e eventos anóxicos; e os ecossistemas marinhos tornaram-se mais diversificados, de modo que as cadeias alimentares eram menos propensas a serem interrompidas.[235][236]
- Fósseis razoavelmente completos são muito raros, a maioria dos organismos extintos é representada apenas por fósseis parciais e fósseis completos são mais raros nas rochas mais antigas. Portanto, os paleontólogos atribuíram erroneamente partes do mesmo organismo a diferentes gêneros, que muitas vezes eram definidos apenas para acomodar essas descobertas—a história de Anomalocaris é um exemplo disso. O risco desse erro é maior para os fósseis mais antigos porque frequentemente são partes diferentes de qualquer organismo vivo e mal conservados. Muitos dos gêneros "supérfluos" são representados por fragmentos que não são encontrados novamente e os gêneros "supérfluos" parecem se extinguir muito rapidamente.[234]

Biodiversidade no registro fóssil, que é "... o número de gêneros distintos vivos em um determinado momento; isto é, aqueles cuja primeira ocorrência é anterior e cuja última ocorrência é posterior àquela época"[237] mostra uma tendência diferente: uma tendência bastante rápida subir de 542 para 400 Ma; um ligeiro declínio de 400 para 200 Ma, no qual o evento devastador de extinção Permiano-Triássico é um fator importante; e um rápido aumento de 200 Ma até o presente.[237]
Ver também
editarNotas
Referências
- ↑ a b Pearce, Ben K.D.; Tupper, Andrew S.; Pudritz, Ralph E.; Higgs, Paul G. (1 de março de 2018). «Constraining the Time Interval for the Origin of Life on Earth». Astrobiology. 18 3 ed. pp. 343–364. Bibcode:2018AsBio..18..343P. ISSN 1531-1074. PMID 29570409. arXiv:1808.09460
 . doi:10.1089/ast.2017.1674
. doi:10.1089/ast.2017.1674
- ↑ a b Rosing, Minik T. (29 de janeiro de 1999). «13C-Depleted Carbon Microparticles in >3700-Ma Sea-Floor Sedimentary Rocks from West Greenland». Science. 283 5402 ed. pp. 674–676. Bibcode:1999Sci...283..674R. ISSN 0036-8075. PMID 9924024. doi:10.1126/science.283.5402.674
- ↑ a b Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T. (Janeiro de 2014). «Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks». Nature Geoscience. 7 1 ed. pp. 25–28. Bibcode:2014NatGe...7...25O. ISSN 1752-0894. doi:10.1038/ngeo2025
- ↑ Papineau, Dominic; De Gregorio, Bradley T.; Cody, George D.; O'Neil, J.; Steele, A.; Stroud, R. M.; Fogel, M. L. (Junho de 2011). «Young poorly crystalline graphite in the >3.8-Gyr-old Nuvvuagittuq banded iron formation». Nature Geoscience. 4 6 ed. pp. 376–379. Bibcode:2011NatGe...4..376P. ISSN 1752-0894. doi:10.1038/ngeo1155
- ↑ Bell, Elizabeth A.; Boehnke, Patrick; Harrison, T. Mark; Mao, Wendy L. (24 de novembro de 2015). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon» (PDF). Proceedings of the National Academy of Sciences. 112 47 ed. pp. 14518–14521. Bibcode:2015PNAS..11214518B. ISSN 0027-8424. PMC 4664351 . PMID 26483481. doi:10.1073/pnas.1517557112. Consultado em 14 de fevereiro de 2020. Cópia arquivada (PDF) em 13 de fevereiro de 2020
- ↑ Nemchin, Alexander A.; Whitehouse, Martin J.; Menneken, Martina; Geisler, Thorsten; Pidgeon, Robert T.; Wilde, Simon A. (3 de julho de 2008). «A light carbon reservoir recorded in zircon-hosted diamond from the Jack Hills». Nature. 454 7200 ed. pp. 92–95. Bibcode:2008Natur.454...92N. ISSN 0028-0836. PMID 18596808. doi:10.1038/nature07102
- ↑ Futuyma, Douglas Joel (2005). Evolution. Sunderland, Massachusetts: Sinuer Associates, Inc. ISBN 0-87893-187-2
- ↑ Dybas, Cheryl; Fryling, Kevin (2 de maio de 2016). «Researchers find that Earth may be home to 1 trillion species». Alexandria, VA: National Science Foundation. News Release 16-052. Consultado em 11 de dezembro de 2016. Cópia arquivada em 4 de maio de 2016
- Locey, Kenneth J.; Lennon, Jay T. (24 de maio de 2016). «Scaling laws predict global microbial diversity». Proc. Natl. Acad. Sci. U.S.A. 113 21 ed. pp. 5970–5975. Bibcode:2016PNAS..113.5970L. ISSN 0027-8424. PMC 4889364 . PMID 27140646. doi:10.1073/pnas.1521291113
- Locey, Kenneth J.; Lennon, Jay T. (24 de maio de 2016). «Scaling laws predict global microbial diversity». Proc. Natl. Acad. Sci. U.S.A. 113 21 ed. pp. 5970–5975. Bibcode:2016PNAS..113.5970L. ISSN 0027-8424. PMC 4889364
- ↑ Chapman 2009.
- ↑ Novacek, Michael J. (8 de novembro de 2014). «Prehistory's Brilliant Future». Sunday Review. The New York Times. Nova Iorque. ISSN 0362-4331. Consultado em 25 de dezembro de 2014. Cópia arquivada em 10 de novembro de 2014 "A version of this article appears in print on Nov. 9, 2014, Section SR, Page 6 of the New York edition with the headline: Prehistory's Brilliant Future."
- ↑ «Catalogue of Life: 2019 Annual Checklist». Species 2000; Integrated Taxonomic Information System. 2019. Consultado em 16 de fevereiro de 2020
- ↑ McKinney 1997, p. 110.
- ↑ Stearns & Stearns 1999, p. x.
- ↑ Nutman, Allen P.; Bennett, Vickie C.; Friend, Clark R.L.; Van Kranendonk, Martin J.; Chivas, Allan R. (22 de setembro de 2016). «Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures» (PDF). Nature. 537 7621 ed. pp. 535–538. Bibcode:2016Natur.537..535N. ISSN 0028-0836. PMID 27580034. doi:10.1038/nature19355. Consultado em 17 de fevereiro de 2020. Cópia arquivada em 2 de janeiro de 2020
- ↑ Borenstein, Seth (19 de outubro de 2015). «Hints of life on what was thought to be desolate early Earth». Excite. Yonkers, NY: Mindspark Interactive Network. Associated Press. Consultado em 20 de outubro de 2015
- ↑ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; Mao, Wendy L. (19 de outubro de 2015). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon» (PDF). Washington, D.C.: National Academy of Sciences. Proc. Natl. Acad. Sci. U.S.A. 112: 14518–21. ISSN 1091-6490. PMC 4664351 . PMID 26483481. doi:10.1073/pnas.1517557112. Consultado em 20 de outubro de 2015 Early edition, published online before print.
- ↑ a b Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; Slack, John F.; Rittner, Martin; Pirajno, Franco; O'Neil, Jonathan; Little, Crispin T. S. (2 de março de 2017). «Evidence for early life in Earth's oldest hydrothermal vent precipitates» (PDF). Nature. 543 7643 ed. pp. 60–64. Bibcode:2017Natur.543...60D. ISSN 0028-0836. PMID 28252057. doi:10.1038/nature21377. Consultado em 18 de fevereiro de 2020. Cópia arquivada (PDF) em 13 de fevereiro de 2020
- ↑ Zimmer, Carl (1 de março de 2017). «Scientists Say Canadian Bacteria Fossils May Be Earth's Oldest». Matter. The New York Times. Nova Iorque. ISSN 0362-4331. Consultado em 2 de março de 2017. Cópia arquivada em 4 de janeiro de 2020 "A version of this article appears in print on March 2, 2017, Section A, Page 9 of the New York edition with the headline: Artful Squiggles in Rocks May Be Earth's Oldest Fossils."
- ↑ a b c Nisbet, E.G., and Fowler, C.M.R. (7 de dezembro de 1999). «Archaean metabolic evolution of microbial mats». Proceedings of the Royal Society: Biology. 266 (1436). 2375 páginas. PMC 1690475 . doi:10.1098/rspb.1999.0934 - resumo com ligação ao conteúdo completo grátis (PDF)
- ↑ Anbar, Ariel D.; Yun (28 de setembro de 2007). «A Whiff of Oxygen Before the Great Oxidation Event?». Science (em inglês). 317 (5846): 1903-1906. ISSN 0036-8075. PMID 17901330. doi:10.1126/science.1140325
- ↑ Bonner, J.T. (1998) The origins of multicellularity. Integr. Biol. 1, 27–36
- ↑ "Os fósseis mais antigos revelam a evolução de plantas não-vasculares até meados ou fins do Período Ordovicíco (~450-440 m.a.a.) com base em esporos fósseis" (em inglês no original) "The oldest fossils reveal evolution of non-vascular plants by the middle to late Ordovician Period (~450-440 m.y.a.) on the basis of fossil spores" Transition of plants to land Arquivado em 2 de novembro de 2013, no Wayback Machine.
- ↑ Algeo, T.J. (1998). «Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events». Philosophical Transactions of the Royal Society B: Biological Sciences. 353 (1365): 113–130. doi:10.1098/rstb.1998.0195
- ↑ «Metazoa: Fossil Record»
- ↑ a b Shu; et al. (4 de novembro de 1999). «Lower Cambrian vertebrates from south China». Nature. 402: 42–46. doi:10.1038/46965
- ↑ Hoyt, Donald F. (1997). «Synapsid Reptiles». Consultado em 1 de junho de 2010. Arquivado do original em 20 de maio de 2009
- ↑ Barry, Patrick L. (28 de janeiro de 2002). «The Great Dying». Science@NASA. Science and Technology Directorate, Marshall Space Flight Center, NASA. Consultado em 26 de março de 2009. Arquivado do original em 16 de fevereiro de 2012
- ↑ Tanner LH, Lucas SG & Chapman MG (2004). «Assessing the record and causes of Late Triassic extinctions» (PDF). Earth-Science Reviews. 65 (1-2): 103–139. doi:10.1016/S0012-8252(03)00082-5. Consultado em 22 de outubro de 2007. Arquivado do original (PDF) em 25 de outubro de 2007
- ↑ Benton, M.J. (2004). Vertebrate Paleontology. [S.l.]: Blackwell Publishers. xii–452. ISBN 0-632-05614-2
- ↑ «Amniota - Palaeos»
- ↑ Fastovsky DE, Sheehan PM (2005). «The extinction of the dinosaurs in North America». GSA Today. 15 (3): 4–10. doi:10.1130/1052-5173(2005)015<4:TEOTDI>2.0.CO;2. Consultado em 18 de maio de 2007. Arquivado do original em 9 de dezembro de 2011
- ↑ «Dinosaur Extinction Spurred Rise of Modern Mammals». News.nationalgeographic.com. Consultado em 8 de março de 2009
- ↑ Van Valkenburgh, B. (1999). «Major patterns in the history of carnivorous mammals». Annual Review of Earth and Planetary Sciences. 26: 463–493. doi:10.1146/annurev.earth.27.1.463
- ↑ Erwin, Douglas H. (9 de novembro de 2015). «Early metazoan life: divergence, environment and ecology» 20150036 ed. Phil. Trans. R. Soc. B. 370. doi:10.1098/rstb.2015.0036. Consultado em 7 de janeiro de 2016
- ↑ a b El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andrey; Macchiarelli, Reberto; Mazurier, Arnaud; Hammarlund, Emma U.; Boulvais, Philippe; Dupuy, Jean-Jacques (1 de julho de 2010). «Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago» 7302 ed. Londres: Nature Publishing Group. Nature. 466: 100–104. Bibcode:2010Natur.466..100A. ISSN 0028-0836. PMID 20596019. doi:10.1038/nature09166
- ↑ a b
- Dalrymple, G.B. (1991). The Age of the Earth. California: Stanford University Press. ISBN 0-8047-1569-6
- Newman, W.L. (2007). «Age of the Earth». Publications Services, USGS. Consultado em 29 de agosto de 2008
- Dalrymple, G.B. (2001). «The age of the Earth in the twentieth century: a problem (mostly) solved». Geological Society, London, Special Publications. 190 (1): 205–221. doi:10.1144/GSL.SP.2001.190.01.14. Consultado em 20 de setembro de 2007
- ↑ Galimov, E.M. and Krivtsov, A.M. (2005). «Origin of the Earth-Moon System». J. Earth Syst. Sci. 114 (6): 593–600. Bibcode:2005JESS..114..593G. doi:10.1007/BF02715942 [1]
- ↑ a b Cohen, B.A., Swindle, T.D. and Kring, D.A. (2000). «Support for the Lunar Cataclysm Hypothesis from Lunar Meteorite Impact Melt Ages». Science. 290 (5497): 1754–1756. Bibcode:2000Sci...290.1754C. PMID 11099411. doi:10.1126/science.290.5497.1754. Consultado em 31 de agosto de 2008
- ↑ * «Early Earth Likely Had Continents And Was Habitable». University of Colorado. 17 de novembro de 2005. Consultado em 11 de janeiro de 2009
- Cavosie, A.J., Valley, J.W., Wilde, S. A. and the Edinburgh Ion Microprobe Facility (15 de julho de 2005). «Magmatic δ18O in 4400-3900 Ma detrital zircons: A record of the alteration and recycling of crust in the Early Archean». Earth and Planetary Science Letters. 235 (3–4): 663–681. Bibcode:2005E&PSL.235..663C. doi:10.1016/j.epsl.2005.04.028
- ↑ Britt, R.R. (24 de julho de 2002). «Evidence for Ancient Bombardment of Earth». Space.com. Consultado em 15 de abril de 2006
- ↑ Valley, J.W., Peck, W.H., King, E.M. and Wilde, S.A. (2002). «A cool early Earth» (PDF). Geology. 30 (4): 351–354. Bibcode:2002Geo....30..351V. doi:10.1130/0091-7613(2002)030<0351:ACEE>2.0.CO;2. Consultado em 13 de setembro de 2008
- ↑ Dauphas, N., Robert, F. and Marty, B. (2000). «The Late Asteroidal and Cometary Bombardment of Earth as Recorded in Water Deuterium to Protium Ratio». Icarus. 148 (2): 508–512. Bibcode:2000Icar..148..508D. doi:10.1006/icar.2000.6489
- ↑ Scalice, Daniella (20 de maio de 2009). Fletcher, Julie, ed. «Microbial Habitability During the Late Heavy Bombardment». Astrobiology. Mountain View, CA: NASA Astrobiology Program. Consultado em 25 de fevereiro de 2020. Cópia arquivada em 24 de janeiro de 2015
- Abramov, Oleg; Mojzsis, Stephen J. (21 de maio de 2009). «Microbial habitability of the Hadean Earth during the late heavy bombardment». Nature. 459 7245 ed. pp. 419–422. Bibcode:2009Natur.459..419A. ISSN 0028-0836. PMID 19458721. doi:10.1038/nature08015
- ↑ a b Brasier, M., McLoughlin, N., Green, O. and Wacey, D. (2006). «A fresh look at the fossil evidence for early Archaean cellular life» (PDF). Philosophical Transactions of the Royal Society: Biology. 361 (1470): 887–902. PMC 1578727 . PMID 16754605. doi:10.1098/rstb.2006.1835. Consultado em 30 de agosto de 2008
- ↑ * Schopf, J. W. (1993). «Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life». Science. 260 (5108): 640–646. Bibcode:1993Sci...260..640S. PMID 11539831. doi:10.1126/science.260.5108.640. Consultado em 30 de agosto de 2008
- Altermann, W. and Kazmierczak, J. (2003). «Archean microfossils: a reappraisal of early life on Earth». Res Microbiol. 154 (9): 611–7. PMID 14596897. doi:10.1016/j.resmic.2003.08.006
- ↑ Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison, T.M., Nutman, A.P. and Friend, C.R.L. (1996). «Evidence for life on Earth before 3.8 Ga». Nature. 384 (6604): 55–59. Bibcode:1996Natur.384...55M. PMID 8900275. doi:10.1038/384055a0. Consultado em 30 de agosto de 2008
- ↑ Grotzinger, J.P. and Rothman, D.H. (1996). «An abiotic model for stomatolite morphogenesis». Nature. 383 (6599): 423–425. Bibcode:1996Natur.383..423G. doi:10.1038/383423a0
- ↑ * Fedo, C.M. and Whitehouse, M.J. (2002). «Metasomatic Origin of Quartz-Pyroxene Rock, Akilia, Greenland, and Implications for Earth's Earliest Life». Science. 296 (5572): 1448–1452. Bibcode:2002Sci...296.1448F. PMID 12029129. doi:10.1126/science.1070336. Consultado em 30 de agosto de 2008
- Lepland, A., van Zuilen, M.A., Arrhenius, G., Whitehouse, M.J. and Fedo, C.M. (2005). «Questioning the evidence for Earth's earliest life — Akilia revisited». Geology. 33 (1): 77–79. Bibcode:2005Geo....33...77L. doi:10.1130/G20890.1. Consultado em 30 de agosto de 2008
- ↑ Schopf, J. (2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society of London: B Biological Sciences. 361 (1470): 869–85. PMC 1578735 . PMID 16754604. doi:10.1098/rstb.2006.1834
- ↑ Drake, Nadia (1 de março de 2017). «This May Be the Oldest Known Sign of Life on Earth». National Geographic News. Washington, D.C.: National Geographic Society. Consultado em 26 de fevereiro de 2020. Cópia arquivada em 23 de outubro de 2019
- ↑ Ciccarelli, Francesca D.; Doerks, Tobias; von Mering, Christian; Creevey, Christopher J.; Snel, Berend; Bork, Peer (3 de março de 2006). «Toward Automatic Reconstruction of a Highly Resolved Tree of Life» 5765 ed. Washington, D.C.: American Association for the Advancement of Science. Science. 311: 1283–1287. Bibcode:2006Sci...311.1283C. ISSN 0036-8075. PMID 16513982. doi:10.1126/science.1123061
- ↑ Mason, Stephen F. (1984). «Origins of biomolecular handedness» 5981 ed. Londres: Nature Publishing Group. Nature. 311: 19–23. Bibcode:1984Natur.311...19M. ISSN 0028-0836. PMID 6472461. doi:10.1038/311019a0
- ↑ Orgel, Leslie E. (Outubro de 1994). «The Origin of Life on the Earth» (PDF) 4 ed. Stuttgart: Georg von Holtzbrinck Publishing Group. Scientific American. 271: 76–83. ISSN 0036-8733. PMID 7524147. doi:10.1038/scientificamerican1094-76. Consultado em 30 de agosto de 2008. Cópia arquivada em 24 de janeiro de 2001.
- ↑ O'Leary, M.R. (2008). Anaxagoras and the Origin of Panspermia Theory. [S.l.]: iUniverse, Inc. ISBN 0-595-49596-6
- ↑ a b Arrhenius, S. (1903). «The Propagation of Life in Space». Die Umschau. 7 Reimpresso em Goldsmith, D., (ed.). The Quest for Extraterrestrial Life. [S.l.]: University Science Books. ISBN 0-19-855704-3
- ↑ Hoyle, F. and Wickramasinghe, C. (1979). «On the Nature of Interstellar Grains». Astrophysics and Space Science. 66: 77–90. Bibcode:1979Ap&SS..66...77H. doi:10.1007/BF00648361
- ↑ a b Crick, F; Orgel, L.E. (1973). «Directed Panspermia». Icarus. 19 (3): 341–348. Bibcode:1973Icar...19..341C. doi:10.1016/0019-1035(73)90110-3 Parâmetro desconhecido
|author-name-list=ignorado (ajuda) - ↑ a b c Warmflash, D. and Weiss, B. (2005). «Did Life Come From Another World?». Scientific American: 64–71. Consultado em 2 de setembro de 2008
- ↑ Clancy, Brack & Horneck 2005
- ↑ Horneck, Gerda; Klaus, David M.; Mancinelli, Rocco L. (Março de 2010). «Space Microbiology» 1 ed. Washington, D.C.: American Society for Microbiology. Microbiology and Molecular Biology Reviews. 74: 121–156. ISSN 1092-2172. doi:10.1128/mmbr.00016-09. Consultado em 29 de julho de 2013
- ↑ Ker, Than (2007). «Claim of Martian Life Called 'Bogus'». space.com. Consultado em 2 de setembro de 2008
- ↑ Bennett 2008, pp. 82–85
- ↑ Schulze-Makuch, Dirk; Irwin, Louis N. (Abril de 2006). «The prospect of alien life in exotic forms on other worlds». Naturwissenschaften. 93 (4): 155–172. Bibcode:2006NW.....93..155S. ISSN 0028-1042. PMID 16525788. doi:10.1007/s00114-005-0078-6
- ↑ Peretó, Juli (Março de 2005). «Controversies on the origin of life» (PDF). International Microbiology. 8 (1): 23–31. ISSN 1139-6709. PMID 15906258. Consultado em 7 de outubro de 2007. Cópia arquivada (PDF) em 4 de junho de 2007
- ↑ Szathmáry, Eörs (3 de fevereiro de 2005). «In search of the simplest cell». Nature. 433 (7025): 469–470. Bibcode:2005Natur.433..469S. ISSN 0028-0836. PMID 15690023. doi:10.1038/433469a
- ↑ Luisi, Pier Luigi; Ferri, Francesca; Stano, Pasquale (Janeiro de 2006). «Approaches to semi-synthetic minimal cells: a review». Naturwissenschaften. 93 (1): 1–13. Bibcode:2006NW.....93....1L. ISSN 0028-1042. PMID 16292523. doi:10.1007/s00114-005-0056-z
- ↑ Joyce, Gerald F. (11 de julho de 2002). «The antiquity of RNA-based evolution». Nature. 418 (6894): 214–221. Bibcode:2002Natur.418..214J. ISSN 0028-0836. PMID 12110897. doi:10.1038/418214a
- ↑ a b Hoenigsberg, Hugo (30 de dezembro de 2003). «Evolution without speciation but with selection: LUCA, the Last Universal Common Ancestor in Gilbert's RNA world». Genetics and Molecular Research. 2 (4): 366–375. ISSN 1676-5680. PMID 15011140. Consultado em 30 de agosto de 2008. Cópia arquivada em 2 de junho de 2004
- ↑ Trevors, Jack T.; Abel, David L. (Novembro de 2004). «Chance and necessity do not explain the origin of life». Cell Biology International. 28 (11): 729–739. ISSN 1065-6995. PMID 15563395. doi:10.1016/j.cellbi.2004.06.006
- ↑ Forterre, Patrick; Benachenhou-Lahfa, Nadia; Confalonieri, Fabrice; Duguet, Michel; Elie, Christiane; Labedan, Bernard (1992). Adoutte, André; Perasso, Roland, eds. «The nature of the last universal ancestor and the root of the tree of life, still open questions». BioSystems. 28 (1–3): 15–32. ISSN 0303-2647. PMID 1337989. doi:10.1016/0303-2647(92)90004-I Part of a special issue: 9th Meeting of the International Society for Evolutionary Protistology, July 3–7, 1992, Orsay, France.
- ↑ Cech, Thomas R. (11 de agosto de 2000). «The Ribosome Is a Ribozyme». Science. 289 (5481): 878–879. ISSN 0036-8075. PMID 10960319. doi:10.1126/science.289.5481.878
- ↑ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (24 de outubro de 2017). «Origin of the RNA world: The fate of nucleobases in warm little ponds». Proceedings of the National Academy of Sciences. 114 (43): 11327–11332. Bibcode:2017PNAS..11411327P. ISSN 0027-8424. PMC 5664528 . PMID 28973920. arXiv:1710.00434 . doi:10.1073/pnas.1710339114
- ↑ Johnston, Wendy K.; Unrau, Peter J.; Lawrence, Michael S.; Glasner, Margaret E.; Bartel, David P. (18 de maio de 2001). «RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension» (PDF). Science. 292 (5520): 1319–1325. Bibcode:2001Sci...292.1319J. CiteSeerX 10.1.1.70.5439 . ISSN 0036-8075. PMID 11358999. doi:10.1126/science.1060786. Cópia arquivada (PDF) em 9 de setembro de 2006
- ↑ a b Levy, Matthew; Miller, Stanley L. (7 de julho de 1998). «The stability of the RNA bases: Implications for the origin of life». Proc. Natl. Acad. Sci. U.S.A. 95 (14): 7933–7938. Bibcode:1998PNAS...95.7933L. ISSN 0027-8424. PMC 20907 . PMID 9653118. doi:10.1073/pnas.95.14.7933
- Larralde, Rosa; Robertson, Michael P.; Miller, Stanley L. (29 de agosto de 1995). «Rates of decomposition of ribose and other sugars: Implications for chemical evolution». Proc. Natl. Acad. Sci. U.S.A. 92 (18): 8158–8160. Bibcode:1995PNAS...92.8158L. ISSN 0027-8424. PMC 41115 . PMID 7667262. doi:10.1073/pnas.92.18.8158
- Lindahl, Tomas (22 de abril de 1993). «Instability and decay of the primary structure of DNA». Nature. 362 (6422): 709–715. Bibcode:1993Natur.362..709L. ISSN 0028-0836. PMID 8469282. doi:10.1038/362709a0
- Larralde, Rosa; Robertson, Michael P.; Miller, Stanley L. (29 de agosto de 1995). «Rates of decomposition of ribose and other sugars: Implications for chemical evolution». Proc. Natl. Acad. Sci. U.S.A. 92 (18): 8158–8160. Bibcode:1995PNAS...92.8158L. ISSN 0027-8424. PMC 41115
- ↑ Orgel, Leslie E. (17 de novembro de 2000). «A Simpler Nucleic Acid». Science. 290 (5495): 1306–1307. ISSN 0036-8075. PMID 11185405. doi:10.1126/science.290.5495.1306
- ↑ Nelson, Kevin E.; Levy, Matthew; Miller, Stanley L. (11 de abril de 2000). «Peptide nucleic acids rather than RNA may have been the first genetic molecule». Proc. Natl. Acad. Sci. U.S.A. 97 (8): 3868–3871. Bibcode:2000PNAS...97.3868N. ISSN 0027-8424. PMC 18108 . PMID 10760258. doi:10.1073/pnas.97.8.3868
- ↑ Martin, William; Russell, Michael J. (29 de janeiro de 2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Philosophical Transactions of the Royal Society B. 358 (1429): 59–85. ISSN 0962-8436. PMC 1693102 . PMID 12594918. doi:10.1098/rstb.2002.1183
- ↑ Wächtershäuser, Günter (25 de agosto de 2000). «Life as We Don't Know It». Science. 289 (5483): 1307–1308. ISSN 0036-8075. PMID 10979855. doi:10.1126/science.289.5483.1307
- ↑ Trevors, Jack T.; Psenner, Roland (Dezembro de 2001). «From self-assembly of life to present-day bacteria: a possible role for nanocells». FEMS Microbiology Reviews. 25 (5): 573–582. ISSN 0168-6445. PMID 11742692. doi:10.1111/j.1574-6976.2001.tb00592.x
- ↑ Segré, Daniel; Ben-Eli, Dafna; Deamer, David W.; Lancet, Doron (Fevereiro de 2001). «The Lipid World» (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119–145. Bibcode:2001OLEB...31..119S. ISSN 0169-6149. PMID 11296516. doi:10.1023/A:1006746807104. Consultado em 28 de fevereiro de 2020. Cópia arquivada (PDF) em 26 de junho de 2015
- ↑ Cairns-Smith 1968, pp. 57–66
- ↑ Ferris, James P. (Junho de 1999). «Prebiotic Synthesis on Minerals: Bridging the Prebiotic and RNA Worlds». The Biological Bulletin. 196 (3): 311–314. ISSN 0006-3185. JSTOR 1542957. PMID 10390828. doi:10.2307/1542957 "This paper was originally presented at a workshop titled Evolution: A Molecular Point of View."
- ↑ Hanczyc, Martin M.; Fujikawa, Shelly M.; Szostak, Jack W. (24 de outubro de 2003). «Experimental Models of Primitive Cellular Compartments: Encapsulation, Growth, and Division». Science. 302 (5645): 618–622. Bibcode:2003Sci...302..618H. ISSN 0036-8075. PMC 4484575 . PMID 14576428. doi:10.1126/science.1089904
- ↑ Hartman, Hyman (Outubro de 1998). «Photosynthesis and the Origin of Life». Origins of Life and Evolution of Biospheres. 28 (4–6): 512–521. Bibcode:1998OLEB...28..515H. ISSN 0169-6149. PMID 11536891. doi:10.1023/A:1006548904157
- ↑ Krumbein et al. 2003, pp. 1–28
- ↑ a b Risatti, J. Bruno; Capman, William C.; Stahl, David A. (11 de outubro de 1994). «Community structure of a microbial mat: The phylogenetic dimension». Proc. Natl. Acad. Sci. U.S.A. 91 (21): 10173–10177. Bibcode:1994PNAS...9110173R. ISSN 0027-8424. PMC 44980 . PMID 7937858. doi:10.1073/pnas.91.21.10173
- ↑ «Biodiversity rocks». Nature (Editor's summary). 441 (7094). 8 de junho de 2006. ISSN 0028-0836. Consultado em 3 de março de 2020. Cópia arquivada em 19 de junho de 2006
- Awramik, Stanley M. (8 de junho de 2006). «Respect for stromatolites». Nature. 441 (7094): 700–701. ISSN 0028-0836. PMID 16760962. doi:10.1038/441700a
- Allwood, Walter & Kamber et al. 2006
- ↑ Allwood, Abigail C.; Walter, Malcolm R.; Kamber, Balz S.; Marshall, Craig P.; Burch, Ian W. (8 de junho de 2006). «Stromatolite reef from the Early Archaean era of Australia». Nature. 441 (7094): 714–718. Bibcode:2006Natur.441..714A. ISSN 0028-0836. PMID 16760969. doi:10.1038/nature04764
- ↑ Blankenship, Robert E. (1 de janeiro de 2001). «Molecular evidence for the evolution of photosynthesis». Trends in Plant Science. 6 (1): 4–6. PMID 11164357. doi:10.1016/S1360-1385(00)01831-8
- ↑ Hoehler, Tori M.; Bebout, Brad M.; Des Marais, David J. (19 de julho de 2001). «The role of microbial mats in the production of reduced gases on the early Earth». Nature. 412 (6844): 324–327. Bibcode:2001Natur.412..324H. ISSN 0028-0836. PMID 11460161. doi:10.1038/35085554
- ↑ Abele, Doris (7 de novembro de 2002). «Toxic oxygen: The radical life-giver» (PDF). Nature. 420 (6911): 27. Bibcode:2002Natur.420...27A. ISSN 0028-0836. PMID 12422197. doi:10.1038/420027a. Consultado em 3 de março de 2020
- ↑ Westerdahl, Becky B. (2007). «Introduction to Aerobic Respiration». Biological Science 10V (Lecture). Davis, CA: University of California, Davis. Consultado em 14 de julho de 2008. Cópia arquivada em 29 de outubro de 2007
- Textbook used for lecture: Starr, Evers & Starr 2007.
- ↑ Goldblatt, Colin; Lenton, Timothy M.; Watson, Andrew J. (2006). «The Great Oxidation at ~2.4 Ga as a bistability in atmospheric oxygen due to UV shielding by ozone» (PDF). Geophysical Research Abstracts. 8 (770). ISSN 1029-7006. SRef-ID: 1607-7962/gra/EGU06-A-00770. Consultado em 5 de março de 2020. Cópia arquivada (PDF) em 26 de setembro de 2007
- ↑ a b Glansdorff, Nicolas; Ying, Xu; Labedan, Bernard (9 de julho de 2008). «The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner». Biology Direct. 3 (29): 29. ISSN 1745-6150. PMC 2478661 . PMID 18613974. doi:10.1186/1745-6150-3-29
- ↑ a b Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E. (1 de agosto de 1999). «Archean Molecular Fossils and the Early Rise of Eukaryotes». Science. 285 (5430): 1033–1036. Bibcode:1999Sci...285.1033B. CiteSeerX 10.1.1.516.9123 . ISSN 0036-8075. PMID 10446042. doi:10.1126/science.285.5430.1033
- ↑ a b c Hedges, S. Blair; Blair, Jaime E.; Venturi, Maria L.; Shoe, Jason L. (28 de janeiro de 2004). «A molecular timescale of eukaryote evolution and the rise of complex multicellular life». BMC Evolutionary Biology. 4: 2. ISSN 1471-2148. PMC 341452 . PMID 15005799. doi:10.1186/1471-2148-4-2
- ↑ Adl, Sina M.; Simpson, Alastair G.B.; Lane, Christopher E. (Setembro de 2012). «The revised classification of eukaryotes» (PDF). Journal of Eukaryotic Microbiology. 59 (5): 429–514. ISSN 1066-5234. PMC 3483872 . PMID 23020233. doi:10.1111/j.1550-7408.2012.00644.x. Consultado em 6 de março de 2020. Cópia arquivada (PDF) em 16 de dezembro de 2013
- Adl, Sina M.; Simpson, Alastair G.B.; Lane, Christopher E. (Maio–Junho de 2013). «Erratum» (PDF). Journal of Eukaryotic Microbiology. 60 (3): 321. ISSN 1066-5234. doi:10.1111/jeu.12033. Consultado em 6 de março de 2020
- ↑ Burki, Fabien (Maio de 2014). «The Eukaryotic Tree of Life from a Global Phylogenomic Perspective». Cold Spring Harbor Perspectives in Biology. 6 (5): a016147. ISSN 1943-0264. PMC 3996474 . PMID 24789819. doi:10.1101/cshperspect.a016147
- ↑ Margulis 1981
- ↑ Vellai, Tibor; Vida, Gábor (7 de agosto de 1999). «The origin of eukaryotes: the difference between prokaryotic and eukaryotic cells». Proceedings of the Royal Society B. 266 (1428): 1571–1577. ISSN 0962-8452. PMC 1690172 . PMID 10467746. doi:10.1098/rspb.1999.0817
- ↑ Selosse, Marc-André; Abert, Béatrice; Godelle, Bernard (1 de março de 2001). «Reducing the genome size of organelles favours gene transfer to the nucleus». Trends in Ecology & Evolution. 16 (3): 135–141. ISSN 0169-5347. PMID 11179577. doi:10.1016/S0169-5347(00)02084-X
- ↑ Pisani, Davide; Cotton, James A.; McInerney, James O. (Agosto de 2007). «Supertrees Disentangle the Chimerical Origin of Eukaryotic Genomes». Molecular Biology and Evolution. 24 (8): 1752–1760. ISSN 0737-4038. PMID 17504772. doi:10.1093/molbev/msm095
- ↑ Gray, Michael W.; Burger, Gertraud; Lang, B. Franz (5 de março de 1999). «Mitochondrial Evolution». Science. 283 (5407): 1476–1481. Bibcode:1999Sci...283.1476G. ISSN 0036-8075. PMC 3428767 . PMID 10066161. doi:10.1126/science.283.5407.1476
- Gray, Michael W. (Setembro de 2012). «Mitochondrial Evolution». Cold Spring Harbor Perspectives in Biology. 4 (9): a011403. ISSN 1943-0264. PMC 3428767 . PMID 22952398. doi:10.1101/cshperspect.a011403
- Gray, Michael W. (Setembro de 2012). «Mitochondrial Evolution». Cold Spring Harbor Perspectives in Biology. 4 (9): a011403. ISSN 1943-0264. PMC 3428767
- ↑ Rasmussen, Birger; Fletcher, Ian R.; Brocks, Jochen J.; et al. (23 de outubro de 2008). «Reassessing the first appearance of eukaryotes and cyanobacteria». Nature. 455 (7216): 1101–1104. Bibcode:2008Natur.455.1101R. ISSN 0028-0836. PMID 18948954. doi:10.1038/nature07381
- ↑ a b c d e Fedonkin, Mikhail A. (31 de março de 2003). «The origin of the Metazoa in the light of the Proterozoic fossil record» (PDF). Tokyo: Palaeontological Society of Japan. Paleontological Research. 7 (1): 9–41. ISSN 1880-0068. doi:10.2517/prpsj.7.9. Consultado em 2 de setembro de 2008. Cópia arquivada (PDF) em 26 de fevereiro de 2009
- ↑ Tsu-Ming, Han; Runnegar, Bruce (10 de julho de 1992). «Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan». Science. 257 (5067): 232–235. Bibcode:1992Sci...257..232H. ISSN 0036-8075. PMID 1631544. doi:10.1126/science.1631544
- ↑ Javaux, Emmanuelle J.; Knoll, Andrew H.; Walter, Malcolm R. (Julho de 2004). «TEM evidence for eukaryotic diversity in mid-Proterozoic oceans». Geobiology. 2 (3): 121–132. ISSN 1472-4677. doi:10.1111/j.1472-4677.2004.00027.x
- ↑ a b Butterfield, Nicholas J. (Verão de 2005). «Probable Proterozoic fungi». Paleontological Society. Paleobiology. 31 (1): 165–182. ISSN 0094-8373. doi:10.1666/0094-8373(2005)031<0165:PPF>2.0.CO;2. Consultado em 30 de janeiro de 2015
- ↑ Wise, Robert R. (2006). «1. The Diversity of Plastid Form and Function». Advances in Photosynthesis and Respiration. 23. [S.l.]: Springer. pp. 3–26. ISBN 978-1-4020-4060-3. doi:10.1007/978-1-4020-4061-0_1
- ↑ a b c d e Neiman, Maurine; Jokela, Jukka (2010). «Sex: Advantage». Encyclopedia of Life Sciences. Hoboken, NJ: John Wiley & Sons. ISBN 0-470-01617-5. doi:10.1002/9780470015902.a0001716.pub2. Consultado em 21 de janeiro de 2015
- ↑ Holmes & Jobling 1996
- ↑ Christie, Peter J. (Abril de 2001). «Type IV secretion: intercellular transfer of macromolecules by systems ancestrally related to conjugation machines». Hoboken, NJ: Wiley-Blackwell. Molecular Microbiology. 40 (2): 294–305. ISSN 0950-382X. PMID 11309113. doi:10.1046/j.1365-2958.2001.02302.x
- ↑ Michod, Richard E.; Bernstein, Harris; Nedelcu, Aurora M. (Maio de 2008). «Adaptive value of sex in microbial pathogens» (PDF). Amsterdã, Holanda: Elsevier. Infection, Genetics and Evolution. 8 (3): 267–285. ISSN 1567-1348. PMID 18295550. doi:10.1016/j.meegid.2008.01.002
- ↑ Bernstein, Harris; Bernstein, Carol (Julho de 2010). «Evolutionary Origin of Recombination during Meiosis». Oxford: Oxford University Press on behalf of the American Institute of Biological Sciences. BioScience. 60 (7): 498–505. ISSN 0006-3568. doi:10.1525/bio.2010.60.7.5
- ↑ Johnsborg, Ola; Eldholm, Vegard; Håvarstein, Leiv Sigve (Dezembro de 2007). «Natural genetic transformation: prevalence, mechanisms and function». Amsterdã, Holanda: Elsevier for the Pasteur Institute. Research in Microbiology. 158 (10): 767–778. ISSN 0923-2508. PMID 17997281. doi:10.1016/j.resmic.2007.09.004
- ↑ a b Bernstein, Bernstein & Michod 2012, pp. 1–50
- ↑ Ramesh, Marilee A.; Malik, Shehre-Banoo; Logsdon, John M., Jr. (26 de janeiro de 2005). «A Phylogenomic Inventory of Meiotic Genes: Evidence for Sex in Giardia and an Early Eukaryotic Origin of Meiosis» (PDF). Cambridge, MA: Cell Press. Current Biology. 15 (2): 185–191. ISSN 0960-9822. PMID 15668177. doi:10.1016/j.cub.2005.01.003. Consultado em 22 de dezembro de 2008. Cópia arquivada (PDF) em 26 de fevereiro de 2009
- ↑ a b Otto, Sarah P.; Gerstein, Aleeza C. (Agosto de 2006). «Why have sex? The population genetics of sex and recombination». Londres: Portland Press on behalf of the Biochemical Society. Biochemical Society Transactions. 34 (Pt 4): 519–522. ISSN 0300-5127. PMID 16856849. doi:10.1042/BST0340519. Consultado em 22 de dezembro de 2008
- ↑ a b Lin Dong; Shuhai Xiao; Bing Shen; Chuanming Zhou (Janeiro de 2008). «Silicified Horodyskia and Palaeopascichnus from upper Ediacaran cherts in South China: tentative phylogenetic interpretation and implications for evolutionary stasis». Londres: Geological Society of London. Journal of the Geological Society. 165 (1): 367–378. ISSN 0016-7649. doi:10.1144/0016-76492007-074. Consultado em 1 de fevereiro de 2015
- ↑ Hanley, Kathryn A.; Fisher, Robert N.; Case, Ted J. (Junho de 1995). «Lower Mite Infestations in an Asexual Gecko Compared With Its Sexual Ancestors». Hoboken, NJ: John Wiley & Sons on behalf of the Society for the Study of Evolution. Evolution. 49 (3): 418–426. ISSN 0014-3820. doi:10.2307/2410266
- ↑ Parker, Matthew A. (Setembro de 1994). «Pathogens and sex in plants». Dordrecht, Holanda: Kluwer Academic Publishers. Evolutionary Ecology. 8 (5): 560–584. ISSN 0269-7653. doi:10.1007/bf01238258
- ↑ Birdsell & Wills 2003, pp. 27–137
- ↑ Bernstein, Hopf & Michod 1987, pp. 323–370
- ↑ Bell, Graham; Mooers, Arne O. (1997). «Size and complexity among multicellular organisms» (PDF). Hoboken, NJ: John Wiley & Sons on behalf of the Linnean Society of London. Biological Journal of the Linnean Society. 60 (3): 345–363. ISSN 0024-4074. doi:10.1111/j.1095-8312.1997.tb01500.x. Consultado em 2 de fevereiro de 2015
- ↑ Kaiser, Dale (Dezembro de 2001). «Building a multicellular organism». Palo Alto, CA: Annual Reviews. Annual Review of Genetics. 35: 103–123. ISSN 0066-4197. PMID 11700279. doi:10.1146/annurev.genet.35.102401.090145
- ↑ a b Bonner, John Tyler (1998). «The origins of multicellularity». Londres: Royal Society of Chemistry. Integrative Biology. 1 (1): 27–36. ISSN 1757-9694. doi:10.1002/(SICI)1520-6602(1998)1:1<27::AID-INBI4>3.0.CO;2-6. Consultado em 3 de setembro de 2008
- ↑ a b Nakagaki, Toshiyuki; Yamada, Hiroyasu; Tóth, Ágota (28 de setembro de 2000). «Intelligence: Maze-solving by an amoeboid organism». Londres: Nature Publishing Group. Nature. 407 (6803). 470 páginas. ISSN 0028-0836. PMID 11028990. doi:10.1038/35035159. Consultado em 3 de setembro de 2008
- ↑ Koschwanez, John H.; Foster, Kevin R.; Murray, Andrew W. (9 de agosto de 2011). «Sucrose Utilization in Budding Yeast as a Model for the Origin of Undifferentiated Multicellularity». São Francisco, CA: Public Library of Science. PLOS Biology. 9 (8): e1001122. ISSN 1545-7885. doi:10.1371/journal.pbio.1001122. Consultado em 2 de fevereiro de 2015
- ↑ a b c d e Butterfield, Nicholas J. (Verão de 2000). «Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes». Paleontological Society. Paleobiology. 26 (3): 386–404. ISSN 0094-8373. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2. Consultado em 1 de fevereiro de 2015
- ↑ Jin JP; et al. (Julho de 2015). «An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors». Molecular Biology and Evolution. 32 (7). doi:10.1093/molbev/msv058
- ↑ Dickey, Gwyneth (31 de julho de 2010). «Evidence for earlier multicellular life» (PNG). Washington, D.C.: Society for Science & the Public. Science News. 178 (3): 17. ISSN 0036-8423. doi:10.1002/scin.5591780322. Consultado em 2 de fevereiro de 2015
- ↑ Gaidos, Eric J.; Dubuc, Timothy; Dunford, Mike; McAndrew, Patricia; Padilla-Gamiño, Jacqueline; Studer, Benjamin; Weersing, Kim; Stanley, Steven (Dezembro de 2007). «The Precambrian emergence of animal life: a geobiological perspective». Geobiology. 5 (4): 351–373. ISSN 1472-4677. doi:10.1111/j.1472-4669.2007.00125.x
- ↑ Jímenez-Guri, Eva; Philippe, Hervé; Okamura, Beth; et al. (6 de julho de 2007). «Buddenbrockia Is a Cnidarian Worm». Science. 317 (5834): 116–118. Bibcode:2007Sci...317..116J. ISSN 0036-8075. PMID 17615357. doi:10.1126/science.1142024
- ↑ Davidson, Michael W. (26 de maio de 2005). «Animal Cell Structure». Molecular Expressions. Tallahassee, FL: Florida State University. Consultado em 3 de setembro de 2008
- ↑ Saupe, Stephen G. (3 de janeiro de 2004). «Concepts of Biology». Concepts of Biology (BIOL116) (Lecture). St. Joseph, MN: College of Saint Benedict and Saint John's University. Consultado em 3 de setembro de 2008
- ↑ Hinde 2001, pp. 28–57
- ↑ a b Chen, Xiaozheng (27 de novembro de 2019). «Researchers say animal-like embryos preceded animal appearance» (Nota de imprensa). American Association for the Advancement of Science. EurekAlert!. Consultado em 28 de novembro de 2019. Cópia arquivada em 28 de novembro de 2019
- ↑ a b Zimmer, Carl (27 de novembro de 2019). «Is This the First Fossil of an Embryo? - Mysterious 609-million-year-old balls of cells may be the oldest animal embryos — or something else entirely.». Matter. The New York Times. Nova Iorque. ISSN 0362-4331. Consultado em 28 de novembro de 2019
- ↑ Cunningham, John A.; Liu, Alexander G.; Bengtson, Stefan; Donoghue, Philip C. J. (Janeiro de 2017). «The origin of animals: Can molecular clocks and the fossil record be reconciled?». BioEssays. 39 (1): e201600120. ISSN 0265-9247. PMID 27918074. doi:10.1002/bies.201600120
- ↑ Jun-Yuan, Chen; Oliveri, Paola; Feng, Gao; Dornbos, Stephen Q.; Chia-Wei Li; Bottjer, David J.; Davidson, Eric H. (1 de agosto de 2002). «Precambrian Animal Life: Probable Developmental and Adult Cnidarian Forms from Southwest China» (PDF). Developmental Biology. 248 (1): 182–196. ISSN 0012-1606. PMID 12142030. doi:10.1006/dbio.2002.0714. Consultado em 4 de fevereiro de 2015. Cópia arquivada (PDF) em 26 de maio de 2013
- ↑ Grazhdankin, Dima (Junho de 2004). «Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution». Paleobiology. 30 (2): 203–221. ISSN 0094-8373. doi:10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2
- ↑ Seilacher, Adolf (Agosto de 1992). «Vendobionta and Psammocorallia: lost constructions of Precambrian evolution». Journal of the Geological Society. 149 (4): 607–613. Bibcode:1992JGSoc.149..607S. ISSN 0016-7649. doi:10.1144/gsjgs.149.4.0607. Consultado em 4 de fevereiro de 2015
- ↑ Martin, Mark W.; Grazhdankin, Dmitriy V.; Bowring, Samuel A.; Evans, David A. D.; Fedonkin, Mikhail A.; Kirschvink, Joseph L. (5 de maio de 2000). «Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution». Science. 288 (5467): 841–845. Bibcode:2000Sci...288..841M. ISSN 0036-8075. PMID 10797002. doi:10.1126/science.288.5467.841
- ↑ Fedonkin, Mikhail A.; Waggoner, Benjamin M. (28 de agosto de 1997). «The late Precambrian fossil Kimberella is a mollusc-like bilaterian organism». Nature. 388 (6645): 868–871. Bibcode:1997Natur.388..868F. ISSN 0028-0836. doi:10.1038/42242
- ↑ Mooi, Rich; David, Bruno (Dezembro de 1998). «Evolution Within a Bizarre Phylum: Homologies of the First Echinoderms». American Zoologist. 38 (6): 965–974. ISSN 0003-1569. doi:10.1093/icb/38.6.965
- ↑ McMenamin, Mark A. S. (Setembro de 2003). Spriggina is a trilobitoid ecdysozoan. Geoscience Horizons Seattle 2003. Abstracts with Programs. 35. Boulder, CO: Geological Society of America. p. 105. ISSN 0016-7592. OCLC 249088612. Consultado em 24 de novembro de 2007. Cópia arquivada em 12 de abril de 2016 Papel No. 40-2 apresentado no Geological Society of America's 2003 Seattle Annual Meeting (2–5 de novembro de 2003) em 2 de novembro de 2003, na Washington State Convention Center.
- ↑ Jih-Pai, Lin; Gon, Samuel M., III; Gehling, James G.; Babcock, Loren E.; Yuan-Long, Zhao; Xing-Liang, Zhang; Shi-Xue, Hu; Jin-Liang, Yuan; Mei-Yi, Yu; Jin, Peng (2006). «A Parvancorina-like arthropod from the Cambrian of South China». Historical Biology. 18 (1): 33–45. ISSN 0891-2963. doi:10.1080/08912960500508689
- ↑ Butterfield, Nicholas J. (Dezembro de 2006). «Hooking some stem-group 'worms': fossil lophotrochozoans in the Burgess Shale». BioEssays. 28 (12): 1161–1166. ISSN 0265-9247. PMID 17120226. doi:10.1002/bies.20507
- ↑ a b c Bengtson 2004, pp. 67–78
- ↑ Gould 1989, pp. 124–136
- ↑ Gould 1989
- ↑ Budd, Graham E. (Fevereiro de 2003). «The Cambrian Fossil Record and the Origin of the Phyla». Integrative and Comparative Biology. 43 (1): 157–165. ISSN 1540-7063. PMID 21680420. doi:10.1093/icb/43.1.157
- ↑ Budd, Graham E. (Março de 1996). «The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group». Lethaia. 29 (1): 1–14. ISSN 0024-1164. doi:10.1111/j.1502-3931.1996.tb01831.x
- ↑ Marshall, Charles R. (30 de maio de 2006). «Explaining the Cambrian 'Explosion' of Animals». Annual Review of Earth and Planetary Sciences. 34: 355–384. Bibcode:2006AREPS..34..355M. ISSN 1545-4495. doi:10.1146/annurev.earth.33.031504.103001
- ↑ Janvier, Philippe (2001). «Vertebrata (Vertebrates)». Encyclopedia of Life Sciences. Hoboken, NJ: John Wiley & Sons. ISBN 978-0-470-01617-6. doi:10.1038/npg.els.0001531
- ↑ Conway Morris, Simon (2 de agosto de 2003). «Once we were worms». New Scientist. 179 (2406). p. 34. ISSN 0262-4079. Consultado em 5 de setembro de 2008. Cópia arquivada em 25 de julho de 2008
- ↑ Jun-Yuan Chen; Di-Ying Huang; Qing-Qing Peng; Hui-Mei Chi; Xiu-Qiang Wang; Man Feng (8 de julho de 2003). «The first tunicate from the Early Cambrian of South China». Proc. Natl. Acad. Sci. U.S.A. 100 (14): 8314–8318. Bibcode:2003PNAS..100.8314C. ISSN 0027-8424. PMC 166226 . PMID 12835415. doi:10.1073/pnas.1431177100
- ↑ D.-G. Shu; Conway Morris, Simon; J. Han; Z.-F. Zhang; K. Yasui; Janvier, Philippe; L. Chen; X.-L. Zhang; J.-N. Liu; Y. Li; H.-Q. Liu (30 de janeiro de 2003). «Head and backbone of the Early Cambrian vertebrate Haikouichthys». Nature. 421 (6922): 526–529. Bibcode:2003Natur.421..526S. ISSN 0028-0836. PMID 12556891. doi:10.1038/nature01264
- ↑ Sansom, Smith & Smith 2001, pp. 156–171
- ↑ a b Cowen 2000, pp. 120–122
- ↑ a b c Selden 2001, "Terrestrialization of Animals," pp. 71–74
- ↑ a b c Garwood, Russell J.; Edgecombe, Gregory D. (Setembro de 2011). «Early Terrestrial Animals, Evolution, and Uncertainty». Evolution: Education and Outreach. 4 (3): 489–501. ISSN 1936-6434. doi:10.1007/s12052-011-0357-y
- ↑ Battistuzzi, Fabia U.; Feijao, Andreia; Hedges, S. Blair (9 de novembro de 2004). «A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land». BMC Evolutionary Biology. 4: 44. ISSN 1471-2148. PMC 533871 . PMID 15535883. doi:10.1186/1471-2148-4-44
- ↑ Weber, Büdel & Belnap 2016, Chapter 3: "Terrestrial Ecosystems in the Precambrian" by Hugo Beraldi-Campesi and Gregory J. Retallack, pp. 37–54. doi:10.1007/978-3-319-30214-0_3: "...terrestrial ecosystems were indeed present, full of life, and functional since the Archean."
- ↑ a b c d e f g Shear 2000, "The Early Development of Terrestrial Ecosystems," pp. 169–184
- ↑ Smith, Deborah (10 de maio de 2017). «Oldest evidence of life on land found in 3.48 billion-year-old Australian rocks» (Nota de imprensa). Sydney, Austrália: UNSW Media. Consultado em 14 de julho de 2020
- ↑ Djokic, Tara; Van Kranendonk, Martin J.; Campbell, Kathleen A.; Walter, Malcolm R.; Ward, Colin R. (9 de maio de 2017). «Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits». Nature Communications. 8: 15263. Bibcode:2017NatCo...815263D. ISSN 2041-1723. PMC 5436104 . PMID 28486437. doi:10.1038/ncomms15263
- ↑ Homann, Martin; Sansjofre, Pierre; Van Zuilen, Mark (Setembro de 2018). «Microbial life and biogeochemical cycling on land 3,220 million years ago» (PDF). Nature Geoscience. 11 (9): 665–671. Bibcode:2018NatGe..11..665H. ISSN 1752-0894. doi:10.1038/s41561-018-0190-9
- ↑ Zimmer, Carl (22 de maio de 2019). «A Billion-Year-Old Fungus May Hold Clues to Life's Arrival on Land». Matter. The New York Times. Nova Iorque. ISSN 0362-4331. Consultado em 23 de maio de 2019. Cópia arquivada em 23 de maio de 2019 "A version of this article appears in print on May 28, 2019, Section D, Page 3 of the New York edition with the headline: Finding a Clue to Life's Arrival."
- ↑ Loron, Corentin C.; François, Camille; Rainbird, Robert H.; Turner, Elizabeth C.; Borensztajn, Stephan; Javaux, Emmanuelle J. (13 de junho de 2019). «Early fungi from the Proterozoic era in Arctic Canada». Nature. 570 (7760): 232–235. Bibcode:2019Natur.570..232L. ISSN 0028-0836. PMID 31118507. doi:10.1038/s41586-019-1217-0
- ↑ Timmer, John (22 de maio de 2019). «Billion-year-old fossils may be early fungus». Ars Technica. Consultado em 23 de maio de 2019. Cópia arquivada em 22 de maio de 2019
- ↑ a b Venturi, Sebastiano (Setembro de 2011). «Evolutionary Significance of Iodine». Current Chemical Biology. 5 (3): 155–162. ISSN 2212-7968. doi:10.2174/187231311796765012
- ↑ Crockford, Susan J. (Agosto de 2009). «Evolutionary roots of iodine and thyroid hormones in cell-cell signaling». Integrative and Comparative Biology. 49 (2): 155–166. ISSN 1557-7023. PMID 21669854. doi:10.1093/icb/icp053
- ↑ Venturi, Sebastiano; Donati, Francesco M.; Venturi, Alessandro; et al. (Agosto de 2000). «Environmental Iodine Deficiency: A Challenge to the Evolution of Terrestrial Life?». Thyroid. 10 (8): 727–729. ISSN 1050-7256. PMID 11014322. doi:10.1089/10507250050137851
- ↑ Küpper, Frithjof C.; Carpenter, Lucy J.; McFiggans, Gordon B.; Palmer, Carl J.; Waite, Tim J.; Boneberg, Eva-Maria; Woitsch, Sonja; Weiller, Markus; Abela, Rafael (13 de maio de 2008). «Iodide accumulation provides kelp with an inorganic antioxidant impacting atmospheric chemistry». Proc. Natl. Acad. Sci. U.S.A. 105 (19): 6954–6958. Bibcode:2008PNAS..105.6954K. ISSN 0027-8424. PMC 2383960 . PMID 18458346. doi:10.1073/pnas.0709959105
- ↑ Lidmar-Bergström, Karna; Bonow, Johan M.; Japsen, Peter (Janeiro de 2013). «Stratigraphic Landscape Analysis and geomorphological paradigms: Scandinavia as an example of Phanerozoic uplift and subsidence». Global and Planetary Change. 100: 153–171. Bibcode:2013GPC...100..153L. ISSN 0921-8181. doi:10.1016/j.gloplacha.2012.10.015
- ↑ Lidmar-Bergström, Karna (Dezembro de 1993). «Denudation surfaces and tectonics in the southernmost part of the Baltic Shield». Precambrian Research. 64 (1–4): 337–345. Bibcode:1993PreR...64..337L. ISSN 0301-9268. doi:10.1016/0301-9268(93)90086-H
- ↑ a b Hawksworth, David L. (2002). «Lichens». Encyclopedia of Life Sciences. Hoboken, NJ: John Wiley & Sons. ISBN 978-0-470-01617-6. doi:10.1038/npg.els.0000368
- ↑ Retallack, Gregory J.; Feakes, Carolyn R. (2 de janeiro de 1987). «Trace Fossil Evidence for Late Ordovician Animals on Land». Science. 235 (4784): 61–63. Bibcode:1987Sci...235...61R. ISSN 0036-8075. PMID 17769314. doi:10.1126/science.235.4784.61
- ↑ a b Kenrick, Paul; Crane, Peter R. (4 de setembro de 1997). «The origin and early evolution of plants on land» (PDF). Nature. 389 (6646): 33–39. Bibcode:1997Natur.389...33K. ISSN 0028-0836. doi:10.1038/37918. Consultado em 10 de fevereiro de 2015
- ↑ Scheckler 2001, "Afforestation—the First Forests," pp. 67–70
- ↑ The phrase "Late Devonian wood crisis" is used at «Tetrapoda: Acanthostega». Palaeos. Consultado em 10 de fevereiro de 2015
- ↑ a b Algeo, Thomas J.; Scheckler, Stephen E. (29 de janeiro de 1998). «Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events». Philosophical Transactions of the Royal Society B. 353 (1365): 113–130. ISSN 0962-8436. PMC 1692181 . doi:10.1098/rstb.1998.0195
- ↑ Taylor, Thomas N.; Osborn, Jeffrey M. (Fevereiro de 1996). «The importance of fungi in shaping the paleoecosystem». Review of Palaeobotany and Palynology. 90 (3–4): 249–262. ISSN 0034-6667. doi:10.1016/0034-6667(95)00086-0
- ↑ Wilson, Heather M.; Anderson, Lyall I. (Janeiro de 2004). «Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland». Journal of Paleontology. 78 (1): 169–184. ISSN 0022-3360. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2
- ↑ Selden, Paul; Read, Helen J. (2008). «The Oldest Land Animals: Silurian Millipedes from Scotland» (PDF). Bulletin of the British Myriapod and Isopod Group. 23: 36–37. ISSN 0267-2154. Consultado em 21 de julho de 2015
- ↑ Shear, William A.; Edgecombe, Gregory D. (Março–Maio de 2010). «The geological record and phylogeny of the Myriapoda». Arthropod Structure & Development. 39 (2–3): 174–190. ISSN 1467-8039. PMID 19944188. doi:10.1016/j.asd.2009.11.002
- ↑ MacNaughton, Robert B.; Cole, Jennifer M.; Dalrymple, Robert W.; Braddy, Simon J.; Briggs, Derek E. G.; Lukie, Terrence D. (Maio de 2002). «First steps on land: Arthropod trackways in Cambrian-Ordovician eolian sandstone, southeastern Ontario, Canada». Geology. 30 (5): 391–394. Bibcode:2002Geo....30..391M. ISSN 0091-7613. doi:10.1130/0091-7613(2002)030<0391:FSOLAT>2.0.CO;2
- ↑ Vaccari, N. Emilio; Edgecombe, Gregory D.; Escudero, C. (29 de julho de 2004). «Cambrian origins and affinities of an enigmatic fossil group of arthropods». Nature. 430 (6999): 554–557. Bibcode:2004Natur.430..554V. ISSN 0028-0836. PMID 15282604. doi:10.1038/nature02705
- ↑ Buatois, Luis A.; Mangano, M. Gabriela; Genise, Jorge F.; Taylor, Thomas N. (Junho de 1998). «The Ichnologic Record of the Continental Invertebrate Invasion: Evolutionary Trends in Environmental Expansion, Ecospace Utilization, and Behavioral Complexity». PALAIOS. 13 (3): 217–240. Bibcode:1998Palai..13..217B. ISSN 0883-1351. JSTOR 3515447. doi:10.2307/3515447
- ↑ Cowen 2000, p. 126
- ↑ Grimaldi & Engel 2005, pp. 155–160
- ↑ Grimaldi & Engel 2005, p. 12
- ↑ a b c Clack, Jennifer A. (Dezembro de 2005). «Getting a Leg Up on Land». Scientific American. 293 (6). pp. 100–107. Bibcode:2005SciAm.293f.100C. ISSN 0036-8733. PMID 16323697. doi:10.1038/scientificamerican1205-100
- ↑ a b Ahlberg, Per E.; Milner, Andrew R. (7 de abril de 1994). «The origin and early diversification of tetrapods». Nature. 368 (6471): 507–514. Bibcode:1994Natur.368..507A. ISSN 0028-0836. doi:10.1038/368507a0
- ↑ Gordon, Malcolm S.; Graham, Jeffrey B.; Wang, Tobias (Setembro–Outubro 2004). «Introduction to the Special Collection: Revisiting the Vertebrate Invasion of the Land». Physiological and Biochemical Zoology. 77 (5): 697–699. ISSN 1522-2152. doi:10.1086/425182
- ↑ Venturi e Bégin 2010, "Thyroid Hormone, Iodine and Human Brain Evolution," pp. 105–124
- ↑ Daeschler, Edward B.; Shubin, Neil H.; Jenkins, Farish A., Jr. (6 de abril de 2006). «A Devonian tetrapod-like fish and the evolution of the tetrapod body plan» (PDF). Nature. 440 (7085): 757–763. Bibcode:2006Natur.440..757D. ISSN 0028-0836. PMID 16598249. doi:10.1038/nature04639. Consultado em 26 de julho de 2020
- ↑ deBraga, Michael; Rieppel, Olivier (Julho de 1997). «Reptile phylogeny and the interrelationships of turtles». Zoological Journal of the Linnean Society. 120 (3): 281–354. ISSN 0024-4082. doi:10.1111/j.1096-3642.1997.tb01280.x
- ↑ a b Benton, Michael J.; Donoghue, Philip C. J. (Janeiro de 2007). «Paleontological Evidence to Date the Tree of Life». Molecular Biology and Evolution. 24 (1): 26–53. ISSN 0737-4038. PMID 17047029. doi:10.1093/molbev/msl150
- ↑ a b Benton, Michael J. (Maio de 1990). «Phylogeny of the major tetrapod groups: Morphological data and divergence dates». Journal of Molecular Evolution. 30 (5): 409–424. Bibcode:1990JMolE..30..409B. ISSN 0022-2844. PMID 2111854. doi:10.1007/BF02101113
- ↑ Sidor, Christian A.; O'Keefe, F. Robin; Damiani, Ross; Steyer, J. Sébastien; Smith, Roger M. H.; Larsson, Hans C. E.; Sereno, Paul C.; Ide, Oumarou; Maga, Abdoulaye (14 de abril de 2005). «Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea». Nature. 434 (7035): 886–889. Bibcode:2005Natur.434..886S. ISSN 0028-0836. PMID 15829962. doi:10.1038/nature03393
- ↑ Smith, Roger; Botha, Jennifer (Setembro–Outubro de 2005). «The recovery of terrestrial vertebrate diversity in the South African Karoo Basin after the end-Permian extinction». Comptes Rendus Palevol. 4 (6–7): 623–636. ISSN 1631-0683. doi:10.1016/j.crpv.2005.07.005
- ↑ Benton 2005
- ↑ Sahney, Sarda; Benton, Michael J. (7 de abril de 2008). «Recovery from the most profound mass extinction of all time». Proceedings of the Royal Society B. 275 (1636): 759–765. ISSN 0962-8452. PMC 2596898 . PMID 18198148. doi:10.1098/rspb.2007.1370
- ↑ Gauthier et al. 1989, p. 345
- ↑ Benton, Michael J. (Março de 1983). «Dinosaur Success in the Triassic: A Noncompetitive Ecological Model» (PDF). The Quarterly Review of Biology. 58 (1): 29–55. ISSN 0033-5770. JSTOR 2828101. doi:10.1086/413056. Consultado em 8 de setembro de 2008. Cópia arquivada (PDF) em 11 de setembro de 2008
- ↑ a b Padian 2004, pp. 210–231
- ↑ Hou, Lian-hai; Zhou, Zhonghe; Martin, Larry D.; Feduccia, Alan (19 de outubro de 1995). «A beaked bird from the Jurassic of China». Nature. 377 (6550): 616–618. Bibcode:1995Natur.377..616H. ISSN 0028-0836. doi:10.1038/377616a0
- ↑ Clarke, Julia A.; Zhou, Zhonghe; Zhang, Fucheng (Março de 2006). «Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui». Journal of Anatomy. 208 (3): 287–308. ISSN 1469-7580. PMC 2100246 . PMID 16533313. doi:10.1111/j.1469-7580.2006.00534.x
- ↑ Ruben, John A.; Jones, Terry D. (Agosto de 2000). «Selective Factors Associated with the Origin of Fur and Feathers». American Zoologist. 40 (4): 585–596. ISSN 0003-1569. doi:10.1093/icb/40.4.585
- ↑ Zhe-Xi, Luo; Crompton, Alfred W.; Ai-Lin, Sun (25 de maio de 2001). «A New Mammaliaform from the Early Jurassic and Evolution of Mammalian Characteristics». Science. 292 (5521): 1535–1540. Bibcode:2001Sci...292.1535L. ISSN 0036-8075. PMID 11375489. doi:10.1126/science.1058476
- ↑ Cifelli, Richard L. (Novembro de 2001). «Early mammalian radiations». Journal of Paleontology. 75 (6): 1214–1226. ISSN 0022-3360. doi:10.1666/0022-3360(2001)075<1214:EMR>2.0.CO;2. Consultado em 16 de fevereiro de 2015
- ↑ Flynn, John J.; Parrish, J. Michael; Rakotosamimanana, Berthe; Simpson, William F.; Wyss, André R. (2 de setembro de 1999). «A Middle Jurassic mammal from Madagascar». Nature. 401 (6748): 57–60. Bibcode:1999Natur.401...57F. ISSN 0028-0836. doi:10.1038/43420
- ↑ MacLeod, Norman; Rawson, Peter F.; Forey, Peter L.; Banner, Frederick T.; Boudagher-Fadel, Marcelle K.; Bown, Paul R.; Burnett, Jackie A.; Chambers, Paul; Culver, Stephen; Evans, Susan E.; Jeffery, Charlotte; Kaminski, Michael A.; Lord, Allan R.; Milner, Angela C.; Milner, Andrew R.; Morris, Noel; Owen, Ellis; Rosen, Brian R.; Smith, Andrew B.; Taylor, Paul D.; Urquhart, Elspeth; Young, Jeremy R. (Abril de 1997). «The Cretaceous–Tertiary biotic transition». Journal of the Geological Society. 154 (2): 265–292. Bibcode:1997JGSoc.154..265M. ISSN 0016-7649. doi:10.1144/gsjgs.154.2.0265. Consultado em 16 de fevereiro de 2015
- ↑ Alroy, John (Março de 1999). «The Fossil Record of North American Mammals: Evidence for a Paleocene Evolutionary Radiation». Systematic Biology. 48 (1): 107–118. ISSN 1063-5157. PMID 12078635. doi:10.1080/106351599260472
- ↑ Archibald, J. David; Deutschman, Douglas H. (Junho de 2001). «Quantitative Analysis of the Timing of the Origin and Diversification of Extant Placental Orders» (PDF). Journal of Mammalian Evolution. 8 (2): 107–124. ISSN 1064-7554. doi:10.1023/A:1011317930838. Consultado em 16 de fevereiro de 2015. Cópia arquivada (PDF) em 9 de julho de 2009
- ↑ Simmons, Nancy B.; Seymour, Kevin L.; Habersetzer, Jörg; Gunnell, Gregg F. (14 de fevereiro de 2008). «Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation». Nature. 451 (7180): 818–821. Bibcode:2008Natur.451..818S. ISSN 0028-0836. PMID 18270539. doi:10.1038/nature06549. hdl:2027.42/62816
- ↑ Thewissen, Madar & Hussain 1996
- ↑ a b c d Crane, Friis e Pedersen 2000, "The Origin and Early Diversification of Angiosperms," pp. 233–250
- ↑ a b c d Crepet, William L. (21 de novembro de 2000). «Progress in understanding angiosperm history, success, and relationships: Darwin's abominably 'perplexing phenomenon'». Proc. Natl. Acad. Sci. U.S.A. 97 (24): 12939–12941. Bibcode:2000PNAS...9712939C. ISSN 0027-8424. PMC 34068 . PMID 11087846. doi:10.1073/pnas.97.24.12939
- ↑ «evolution: plant timeline». Encyclopædia Britannica Online. Encyclopædia Britannica, Inc. 1996. ISBN 9780691185507. OCLC 42796406. Consultado em 23 de outubro de 2020. Cópia arquivada em 27 de março de 2015
- ↑ a b Wilson, Edward O.; Hölldobler, Bert (20 de setembro de 2005). «Eusociality: Origin and consequences». Proc. Natl. Acad. Sci. U.S.A. 102 (38): 13367–13371. Bibcode:2005PNAS..10213367W. ISSN 0027-8424. PMC 1224642 . PMID 16157878. doi:10.1073/pnas.0505858102
- ↑ Hughes, William O. H.; Oldroyd, Benjamin P.; Beekman, Madeleine; Ratnieks, Francis L. W. (30 de maio de 2008). «Ancestral Monogamy Shows Kin Selection Is Key to the Evolution of Eusociality». Science. 320 (5880): 1213–1216. Bibcode:2008Sci...320.1213H. ISSN 0036-8075. PMID 18511689. doi:10.1126/science.1156108
- ↑ Lovegrove, Barry G. (Janeiro de 1991). «The evolution of eusociality in molerats (Bathyergidae): a question of risks, numbers, and costs». Behavioral Ecology and Sociobiology. 28 (1): 37–45. ISSN 0340-5443. doi:10.1007/BF00172137
- ↑ Labandeira & Eble 1999
- ↑ Montgomery, Stephen (2009). «Darwin & The Descent of Man». Charles Darwin & Evolution. Cambridge: Christ's College. Consultado em 19 de novembro de 2020. Cópia arquivada em 20 de junho de 2019
- ↑ Brunet, Michel; Guy, Franck; Pilbeam, David; Mackaye, Hassane Taisso; Likius, Andossa; Ahounta, Djimdoumalbaye; Beauvilain, Alain; Blondel, Cécile (11 de julho de 2002). «A new hominid from the Upper Miocene of Chad, Central Africa». Nature. 418 (6894): 145–151. Bibcode:2002Natur.418..145B. ISSN 0028-0836. PMID 12110880. doi:10.1038/nature00879
- ↑ de Heinzelin, Jean; Clark, J. Desmond; White, Tim; et al. (23 de abril de 1999). «Environment and Behavior of 2.5-Million-Year-Old Bouri Hominids». Science. 284 (5414): 625–629. Bibcode:1999Sci...284..625D. ISSN 0036-8075. PMID 10213682. doi:10.1126/science.284.5414.625
- ↑ De Miguel, Carmen; Henneberg, Maciej (2001). «Variation in hominid brain size: How much is due to method?». HOMO: Journal of Comparative Human Biology. 52 (1): 3–58. ISSN 0018-442X. PMID 11515396. doi:10.1078/0018-442X-00019
- ↑ Leakey 1994, pp. 87–89
- ↑ Mellars, Paul (20 de junho de 2006). «Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model». Proc. Natl. Acad. Sci. U.S.A. 103 (25): 9381–9386. Bibcode:2006PNAS..103.9381M. ISSN 0027-8424. PMC 1480416 . PMID 16772383. doi:10.1073/pnas.0510792103
- ↑ Benton 2005a, Chapter 6: "Tetrapods of the Triassic"
- ↑ a b MacLeod, Norman (1999). «Extinction!». FirstScience.com. Consultado em 25 de dezembro de 2020. Cópia arquivada em 19 de agosto de 2000
- ↑ Martin, Ronald E. (Junho de 1995). «Cyclic and secular variation in microfossil biomineralization: clues to the biogeochemical evolution of Phanerozoic oceans». Global and Planetary Change. 11 (1–2): 1–23. Bibcode:1995GPC....11....1M. ISSN 0921-8181. doi:10.1016/0921-8181(94)00011-2
- ↑ Martin, Ronald E. (Junho de 1996). «Secular Increase in Nutrient Levels through the Phanerozoic: Implications for Productivity, Biomass, and Diversity of the Marine Biosphere». PALAIOS. 11 (3): 209–219. Bibcode:1996Palai..11..209M. ISSN 0883-1351. JSTOR 3515230. doi:10.2307/3515230
- ↑ a b Rohde, Robert A.; Muller, Richard A. (10 de março de 2005). «Cycles in fossil diversity» (PDF). Nature. 434 (7030): 208–210. Bibcode:2005Natur.434..208R. ISSN 0028-0836. PMID 15758998. doi:10.1038/nature03339. Consultado em 22 de setembro de 2008
Bibliografia
editar- Benton, Michael J. (2005). Vertebrate Palaeontology 3 ed. Malden, MA: Blackwell Science. ISBN 978-0-632-05637-8. LCCN 2003028152. OCLC 53970617
- Briggs, Derek E. G.; Crowther, Peter R., eds. (2001). Palaeobiology II. Prefácio por E. N. K. Clarkson. Malden, MA: Blackwell Science. ISBN 978-0-632-05149-6. LCCN 0632051477. OCLC 43945263
- Cowen, R. (2004). History of Life 4th ed. [S.l.]: Blackwell Publishing Limited
- Cunnane, Stephen C.; Stewart, Kathlyn M., eds. (2010). Environmental Influences on Human Brain Evolution. [S.l.]: John Wiley & Sons. ISBN 978-0-470-45268-4. LCCN 2009047979. OCLC 940843566
- Dawkins, R. (2004). The Ancestor's Tale, A Pilgrimage to the Dawn of Life. Boston: Houghton Mifflin Company
- Gee, Henry, ed. (2000). Shaking the Tree: Readings from Nature in the History of Life. Chicago, IL: University of Chicago Press. ISBN 978-0-226-28497-2. LCCN 99049796. OCLC 42476104
- Richard Dawkins. (1990). The Selfish Gene. [S.l.]: Oxford University Press
- Smith, John Maynard; Eörs Szathmáry (1997). The Major Transitions in Evolution. Oxfordshire: Oxford University Press
Ligações externas
editarInformação geral
- General information on evolution- Fossil Museum nav.
- Understanding Evolution from University of California, Berkeley
- National Academies Evolution Resources
- Evolution poster- PDF format "tree of life"
- Everything you wanted to know about evolution by New Scientist
- Howstuffworks.com — How Evolution Works
- Synthetic Theory Of Evolution: An Introduction to Modern Evolutionary Concepts and Theories
História do pensamento evolutivo