Os receptores enzimáticos ou receptores ligados a enzimas são o segundo tipo principal de receptores de membrana celular com domínios intracelulares que estão associados a uma enzima. Em alguns casos, o domínio intracelular do próprio receptor é uma enzima. Outros receptores ligados a enzimas têm um pequeno domínio intracelular que interage diretamente com uma enzima.[1] Esses receptores de membrana medeiam as ações de uma ampla variedade de proteínas mediadoras, incluindo fatores de crescimento e citocinas, e hormônios como a insulina e a leptina, cujos efeitos são exercidos principalmente em nível de transcrição gênica. Os receptores enzimáticos desempenham um papel na apoptose, diferenciação celular, divisão celular, crescimento celular, resposta imune, inflamação e reparo de tecidos. A estrutura dessa classe de receptores polipeptídeos é composta por três domínios principais: o domínio de ligação ao ligante, porção extracelular que permite acesso ao ligante; Domínio transmembrana, composto de uma série de aminoácidos hidrofóbicos que amarra o receptor à membrana celular; Domínio de “enzima ativa” intracelular, intrínseco ao receptor ou fortemente ligado ao domínio transmembrana. A maioria dos domínios de “enzimas ativas” são quinases que fosforilam os aminoácidos serina, treonina e tirosina das proteínas. Quando um ligante se liga ao domínio extracelular leva à dimerização de dois receptores vizinhos. Os receptores dimerizados vizinhos se auto-fosforilam e as proteínas de domínio SH2 ligam-se aos receptores fosforilados e são então fosforiladas, permitindo a continuação do sinal, levando eventualmente à transcrição do gene que leva a uma resposta. [2]
Os receptores tirosina-cinase ou as tirosinas-cinase receptoras incluem os receptores de hormônios, como insulina; fatores de crescimento, como EGF, PDGF, NGF, FGF, VEGF; e efrinas. Com exceção do receptor de insulina, que tem cadeias α e β, essas macromoléculas consistem em cadeias polipeptídicas simples com domínios extracelulares grandes ricos em cisteína, domínios transmembrana curtos e uma região intracelular, que contém um ou dois domínios de tirosina-cinase proteica. A ativação dos receptores dos fatores de crescimento possibilita a sobrevivência, a proliferação e a diferenciação das células. A ativação dos receptores de efrinas estimula a angiogênese neuronal e a migração e o direcionamento axonais. O estado inativo dos receptores dos fatores de crescimento é monomérico; o acoplamento do ligante estimula a dimerização do receptor e a fosforilação cruzada dos domínios de cinase em diversas moléculas de tirosina.[3] As tirosina quinases receptoras fosforilam tirosinas específicas em um pequeno conjunto de proteínas de sinalização intracelular. Uma coleção completa de proteínas de sinalização intracelular pode se ligar às fosfotirosinas em tirosina quinases receptoras ativadas (ou em proteínas de acoplamento especiais, como IRS-1) para ajudar a retransmitir o sinal adiante. Algumas proteínas acopladas são enzimas, como a fosfolipase C-γ (PLC-γ), que funciona da mesma maneira que a fosfolipase C-β - ativando a via de sinalização de fosfolipídio inositol. Por meio dessa via, as tirosina quinases receptoras podem aumentar os níveis de Ca2 + citosólico. Com muito mais frequência, esses receptores dependem mais das cadeias de retransmissão das interações proteína-proteína. Por exemplo, outra enzima que se fixa nesses receptores é a tirosina quinase Src citoplasmática, que fosforila outras proteínas sinalizadoras nas tirosinas. Outra ainda é a fosfatidilinositol 3′-quinase (PI 3-quinase), que gera moléculas lipídicas específicas na membrana plasmática para atrair outras proteínas de sinalização. Embora as proteínas de sinalização intracelular que se ligam a fosfotirosinas em tirosina quinases receptoras ativadas e proteínas de docking tenham estruturas e funções variadas, elas geralmente compartilham domínios de ligação de fosfotirosina altamente conservados. Estes podem ser domínios SH2 (para a região de homologia Src, porque foi primeiro encontrada na proteína Src) ou, menos comumente, domínios PTB (para ligação de fosfotirosina). Ao reconhecer tirosinas fosforiladas específicas, esses pequenos domínios servem como módulos que permitem que as proteínas que os contêm se liguem a tirosina quinases receptoras ativadas, bem como a muitas outras proteínas de sinalização intracelular que foram transitoriamente fosforiladas em tirosinas. Muitas proteínas de sinalização também contêm outros módulos de proteína que lhes permitem interagir especificamente com outras proteínas como parte do processo de sinalização. Estes incluem o domínio SH3 (novamente, assim chamado porque foi descoberto pela primeira vez em Src), que se liga a motivos ricos em prolina em proteínas intracelulares.[4] As subclasses desses receptores incluem: Receptores de fator de crescimento epidérmico, receptores de insulina, receptores de fator de crescimento de nervo [NGF] e receptores tipo Toll [TLRs].
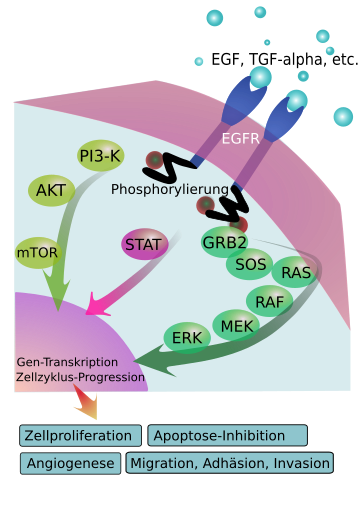
Sinalização da via do EFGRReceptores de fator de crescimento epidérmico [EGFR, ErbB1, HER1]- Esta subclasse RTK é ativada pelo fator de crescimento epidérmico resultando na divisão celular que leva ao crescimento, proliferação e diferenciação celular. Esses receptores são encontrados em níveis anormalmente elevados na superfície de vários tipos de células cancerosas, permitindo que essas células se multipliquem excessivamente.[2] A família RTK do fator de crescimento epidérmico consiste em quatro membros: EGFR (ErbB1, HER1), ErbB2 (HER2, em roedores), ErbB3 (HER3) e ErbB4 (HER4). Estes receptores estruturalmente relacionados são glicoproteínas transmembranares de cadeia simples que consistem em um ectodomínio de ligação ao ligante extracelular, um domínio transmembranar, uma curta seção justamembrana, um domínio tirosina quinase e uma cauda C-terminal contendo tirosina. A ligação do ligante solúvel ao ectodomínio do receptor promove a formação de homo e heterodímero entre os receptores. A dimerização do receptor é essencial para a ativação do domínio tirosina quinase intracelular e fosforilação da cauda C-terminal. Resíduos de fosfotirosina então ativam, diretamente ou através de proteínas adaptadoras, componentes a jusante das vias de sinalização, incluindo Ras / MAPK, PLCγ1 / PKC, PI quinase / Akt e vias STAT. EGFR e ErbB4 podem ser considerados como receptores totalmente funcionais com a capacidade de ligar ligantes e autofosforilar caudas C-terminais através de domínios funcionais de tirosina quinase intracelulares. ErbB2, no entanto, é único por não ter nenhum ligante conhecido, mas é o parceiro de dimerização preferido para outros EGFRs. ErbB3 também é único, pois não tem atividade intrínseca da tirosina quinase, mas pode transduzir sinais por meio da interação com os receptores ativos da quinase, a saber, EGFR, ErbB2 e ErbB4. Embora todos os receptores ErbB tenham sido localizados no núcleo, ErbB4 é notável por uma capacidade convincente de transduzir diretamente sinais extracelulares para o núcleo através da liberação do domínio intracelular por uma clivagem de protease dupla dependente de ligante do receptor. Os estudos cristalográficos dos membros da família EGFR forneceram mais informações sobre a biologia dos receptores e ajudaram a elucidar a base estrutural para as diferenças na função do receptor. O ectodomínio de ligação ao ligante é composto por quatro subdomínios denominados L1 (repetições 1 rica em leucina), CR1 (1 rica em cisteína), L2 e CR2 (alternativamente I - IV). CR1 contém uma alça em gancho β essencial para a função do receptor. Estruturas de cristal de ErbB3 e ErbB4 mostram que, na forma inativa e livre de ligante, o ectodomínio assume uma conformação "amarrada" na qual a alça em gancho beta de CR1 interage com CR2 sequestrando a alça de dimerização. As estruturas do ectodomínio EGFR com EGF ou TGF-α demonstram que a ligação do ligante aos domínios L1 e L2 leva a uma mudança conformacional na qual o receptor assume uma forma estendida que expõe esta alça de dimerização e permite a interação dos ectodomínios do receptor. Um complexo 2: 2 é formado por essa interação na qual os ligantes ficam voltados para fora da face de interação dos dois ectodomínios. Consistente com a incapacidade de identificar um ligante para ErbB2, a estrutura cristalina de um ectodomínio ErbB2 truncado revela que o receptor está bloqueado na conformação estendida pronta para interagir com outros receptores. A conformação estendida abole o potencial local de ligação ao ligante fixando os domínios L1 e L2 em estreita proximidade. Os membros da família EGFR ativam as vias Ras / MAPK, PI (3) K / Akt, PLCγ1 / PKC, STAT e recentemente as vias Par6-atípicas de PKC. A fosforilação da tirosina de EGFR cria sítios de ligação para Grb2 e homologia Src 2 (Shc2), que ativam a via Ras / Raf / MAPK por meio da proliferação celular estimulando a molécula Sos. Outros receptores ErbB também ativam a via Ras / MAPK. A via PI (3) K / Akt é importante para a sobrevivência celular. A ativação de alto nível desta via através da ligação direta da subunidade p85 de PI (3) K ao receptor ativado ocorre com ErbB3 e ErbB4, pois esses receptores contêm locais de ligação a p85. EGFR também pode ativar fracamente a via PI (3) K através da proteína adaptadora Gab-1. O EGFR também é capaz de ativar o PLCγ, levando à ativação da proteína quinase C seguida pela ativação de c-Jun e MAPK, regulando a proliferação celular. STAT3 desempenha um papel importante na manutenção da polaridade e adesão das células epiteliais. A ligação de STAT3 ao EGFR ativado leva à dimerização e translocação de STAT3 para o núcleo para regular a transcrição do gene. Da mesma forma, STAT5 pode se ligar a EGFR e ErbB4. O complexo de Par6-Par3-PKC atípico é importante na organização epitelial. O complexo interage com o ErbB2 ativado, causando a dissociação do Par3 do complexo e resultando na perda da polaridade apical-basal. As células ativadas dessa maneira também apresentam aumento da proliferação e sobrevivência celular, embora os mecanismos subjacentes a todas essas mudanças ainda não tenham sido determinados. [5]
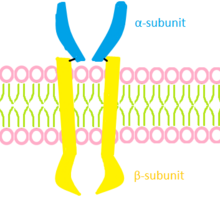
Estrutura de um receptor de insulinaReceptores de insulina- A insulina usa uma classe única de receptores para desencadear o aumento da captação de glicose e aminoácidos e para regular o metabolismo do glicogênio e triglicerídeos na célula. A ativação do receptor em células-alvo específicas dirige um programa complexo de eventos celulares, variando desde transporte em membrana alterado de íons e metabólitos a mudanças na expressão de muitos genes. [6] Uma vez ativado, o receptor de insulina leva a uma cascata de eventos, resultando na expressão de transportadores de glicose (GLUTs) na superfície de uma célula para permitir que ela traga glicose para utilização de energia.[2] Como em todas as quinases, a arquitetura apresenta dois lóbulos estruturalmente distintos, o lóbulo N-terminal e o lóbulo C-terminal, que formam o sítio catalítico da quinase onde o ATP, os cátions bivalentes e o resíduo de tirosina do substrato se unem. Os dois lóbulos são ligados por uma região de ligação que forma uma dobradiça que permite o movimento relativo dos dois lóbulos. Sendo assim esse receptor se apresenta como uma proteína heterotetramérica com atividade quinase, composta por duas subunidades α e duas subunidades β, que atua como uma enzima alostérica na qual a subunidade α inibe a atividade tirosina quinase da subunidade β. A ligação da insulina à subunidade α permite que a subunidade β adquira atividade quinase levando a alteração conformacional e autofosforilação, que aumenta ainda mais a atividade quinase do receptor. [7] Apesar do considerável progresso recente nas estruturas dos domínios extracelular e quinase dos receptores de insulina (e em menor extensão IGF-I), nenhuma estrutura dos receptores completos não ligados e ligados está disponível e, portanto, não se tem os detalhes do mecanismo pelo qual a ligação do ligante extracelular resulta na aproximação e ativação dos domínios da quinase. O que se sabe é que após a ativação do receptor de insulina tirosina quinase por trifosforilação de sua alça de ativação, a quinase fosforila resíduos de tirosina fora do domínio quinase do receptor, que cria sítios de ligação para sinalizar parceiros protéicos contendo SH2 (src-homologia 2) domínios ou domínios PTB (ligação de fosfotirosina). Ao contrário de outros RTKs, os receptores de insulina (e IGF-I) não se ligam a proteínas de sinalização diretamente, mas, em vez disso, ligam-se ao resíduo de domínio de justamembrana fosforilado Tyr 960 (IR - numeração A), uma família de grandes proteínas de ancoragem chamadas IRS (substrato do receptor de insulina ) 1-6, o primeiro dos quais foi clonado em 1991, bem como o adaptador Shc (contendo o domínio de homologia 2 de Src). Estes formam o núcleo para a montagem de uma partícula de transdução de sinal que é o centro de partida das várias cascatas de sinalização intracelulares. Há várias proteínas sinalizadoras envolvidas na insulina. Os três nódulos mais bem estudados são o complexo receptor de insulina / IRS, PI3 quinase (PI3K) e AKT / PKB.A maioria dos efeitos da insulina parece ser mediada por meio da interação de IRS-1 e -2, e Shc, com o receptor de insulina. Outras proteínas de docking como CBL, APS, SH2B, GAB1 and-2 e DOCK1 e-2 foram menos estudadas. As proteínas IRS contêm um domínio de homologia de pleckstrina N-terminal (PH) (que se liga aos fosfolipídios da membrana plasmática) adjacente a um domínio PTB que se liga a um motivo NPXY (Y 960) fosforilado no domínio justamembrana do receptor. As partes central e C-terminal das proteínas IRS contêm até 20 locais de fosforilação potenciais que, quando fosforilados pelo receptor de insulina, se ligam a proteínas de sinalização que contêm domínios SH2.As duas principais vias de sinalização da insulina que emanam do nodo receptor de insulina-IRS são a via fosfatidilinositol 3-quinase (PI3K, uma quinase lipídica) / AKT (também conhecida como PKB ou proteína quinase B) e a via Raf / Ras / MEK / MAPK (proteína quinase ativada por mitogênio, também conhecida como ERK ou quinase regulada por sinal extracelular). A via PI3K é responsável pela maioria dos efeitos metabólicos da insulina e está conectada exclusivamente por IRS, enquanto a via MAPK emana de IRS e Shc e está envolvida na regulação da expressão gênica e, em cooperação com a via PI3K, no controle de crescimento celular ("mitogênese") e diferenciação. [8]
Receptores de fator de crescimento de nervo [NGF]- Essa subclasse de RTK é ativada por fatores de crescimento do nervo, mais especificamente neurotrofinas; uma família de proteínas envolvidas com o desenvolvimento, sobrevivência e função dos neurônios. [2] Os fatores de crescimento são produzidos pelo nosso corpo e têm um papel muito extenso na regulação de muitos processos celulares. A ligação de fatores de crescimento a seus receptores na superfície celular afeta a sobrevivência, proliferação e / ou diferenciação celular. O NGF é outro importante fator de crescimento polipeptídico que funciona para regular o crescimento e a sobrevivência das células nervosas; foi descoberto por Rita Levi-Montalcini e Stanley Cohen na década de 1950. O NGF pertence a uma família de fatores também conhecida como neurotrofinas. Outros membros da família das neurotrofinas incluem fator neurotrófico derivado do cérebro (BDNF), neurotrofina-3 (NT-3) e neurotrofina 4/5 (NT-4/5); todos eles são conhecidos por regular o desenvolvimento e as funções do cérebro. O NGF é formado por clivagem do Pro-NGF, que é a forma de proteína precursora do NGF; no entanto, os papéis do Pro-NGF e NGF não são consistentes; o tratamento com Pro-NGF em neurônios dos gânglios cervicais que expressam ambos os receptores de NGF p75NTR e o receptor de tirosina quinase A (TrkA) leva à morte celular programada, enquanto o tratamento com NGF desses mesmos neurônios resulta em sobrevivência e crescimento axonal. O NGF livre exibe múltiplas ações fisiológicas no sistema nervoso central. O NGF possui efeitos neurotróficos e é crítico para o crescimento e sobrevivência e manutenção dos neurônios. Mais importante, o NGF tem um forte efeito antiapoptótico e, com a privação de NGF, os neurônios exibem uma série de alterações morfológicas e eventualmente sofrem apoptose. O NGF desempenha o seu papel ligando-se aos seus receptores localizados na superfície das células. TrkA é o receptor cataliticamente ativo de alta afinidade para NGF. A ligação de NGF a TrkA leva à fosforilação de TrkA e ativação de seus alvos a jusante, como a proteína quinase B (Akt) ou a proteína quinase 1/2 regulada por sinal extracelular (ERK1 / 2), que eventualmente causa diferenciação neural e prevenção de apoptose. O outro receptor de NGF, p75NTR, é um receptor de baixa afinidade. O papel preciso do p75NTR é complicado e, dependendo do contexto celular, pode promover a sobrevivência celular, a morte celular ou a inibição do crescimento; A afinidade da ligação de NGF ao receptor p75NTR é mais fraca do que a ligação de NGF a TrkA, mas a distribuição de tipo de célula de p75NTR é mais ampla do que a de TrkA; O receptor TrkA é expresso principalmente em neurônios que respondem ao NGF: neurônios sensoriais periféricos, neurônios simpáticos e neurônios colinérgicos do prosencéfalo basal, enquanto o receptor p75NTR é mais amplamente distribuído.Além das células que expressam o receptor TrkA, o receptor p75NTR pode ser detectado em neurônios motores, células de Schwann e células de Purkinje cerebelares. TrkA é o receptor catalítico de alta afinidade para o NGF e medeia os principais efeitos do NGF, que incluem o crescimento celular, a formação e regeneração de neurites e a prevenção da morte celular programada. A ligação de NGF ao receptor TrkA facilita a dimerização do receptor e a autofosforilação de seus resíduos de tirosina. A ativação (fosforilação) do receptor TrkA fornece locais de ancoragem para moléculas efetoras, como Shc que, por sua vez, induz o recrutamento de um complexo de Shc / Grb2, após o qual várias cascatas de sinalização a jusante são iniciadas e propagadas. A fosforilação do receptor TrkA leva à interação de TrkA e fosfatidilinositiol-3 quinase (PI3K). PI3K é ativada e recrutada para a membrana plasmática e leva à produção de fosfoinositídeo 3,4,5-trifosfato e translocação da membrana das proteínas quinases serina / treonina Akt e ativação de Akt. Embora existam outros alvos a jusante de PI3K, a via de sinalização PI3K / Akt é particularmente importante para a sobrevivência neuronal e a síntese de muitas novas proteínas celulares. Outra via de sinalização ativada por NGF é a ativação mediada por Ras da via de proteína quinase ativada por mitógeno (MAPK), que é iniciada por meio de recrutamento e fosforilação de Shc. Ras é uma proteína G associada à membrana; a proteína Ras ativa se liga e fosforila várias proteínas, incluindo o protooncogene Raf. Raf, por sua vez, ativa a MAPK quinase (MEK) e a MEK fosforilada ativa ERK1 / 2. ERK1 / 2 fosforilado pode entrar no núcleo e regular a atividade de muitos fatores de transcrição, incluindo a proteína ELK1 contendo o domínio ETS. ERK1 / 2 também pode fosforilar a quinase S6 ribossomal (S6K), o que leva à fosforilação da proteína de ligação do elemento de resposta ao monofosfato de adenosina cíclica, eventualmente afetando a regulação da expressão de genes indutíveis por NGF e, portanto, contribuindo para a diferenciação neuronal ou crescimento de neuritos . Além das duas vias mencionadas acima, a ativação de TrkA também leva à sobrevivência e ao crescimento de células neuronais por meio da Fosfolipase C gama1 (PLCγ1). O PLCγ1 suporta a ativação da via de sinalização PKC e, portanto, está envolvido na sinalização antimitogênica / mitogênica. O outro receptor de NGF é o p75NTR, mas não é um receptor específico de NGF, pois também se liga a outras neurotrofinas, como NT-3, NT-4/5 e BDNF. O papel do receptor p75NTR é complicado. A ligação de NGF com p75NTR com baixa afinidade leva à apoptose ou sobrevivência celular em diferentes contextos celulares. O p75NTR pode induzir a apoptose in vitro e in vivo. p75NTR ativa Rac GTPase e c-jun ativada N-terminal quinase (JNK), incluindo uma isoforma específica de lesão, JNK3. JNKs estimulam a expressão de genes pró-apoptóticos por meio da transativação de fatores de transcrição específicos, de modo que o p75NTR pode promover a morte celular. No entanto, também aumenta a sobrevivência celular; O tratamento com NGF ativa o fator nuclear κB (NF-κB) por meio de p75NTR e, durante este processo, a ativação de NF-κB mediada por p75NTR aumenta a resposta de sobrevivência dos neurônios sensoriais em desenvolvimento ao fator de crescimento nervoso. NF-κB é um fator de transcrição nuclear que regula a expressão de um grande número de genes que são críticos para a regulação da sobrevivência celular. Assim, em resumo, o receptor p75NTR ativa a sinalização apoptótica através da cascata JNK ou sobrevivência celular através da via NF-κB. O NGF liga-se aos receptores TrkA e p75NTR quando eles coexpressam na membrana celular externa, embora a afinidade do TrkA com o NGF seja muito maior. Estudos in vitro mostraram que os neurônios que coexpressam p75NTR e TrkA respondem a concentrações mais baixas de NGF, o que significa que p75NTR aumenta a capacidade de resposta de TrkA ao NGF. Quando os dois receptores são co-expressos, a taxa de associação de NGF com TrkA aumenta em comparação com as células que expressam TrkA sozinho. O resultado infere que esta interação leva à formação de locais de ligação com maior afinidade para o NGF do que a de qualquer receptor sozinho. As percepções estruturais e mecanísticas das interações do NGF com os receptores TrkA e p75NTR indicam que o NGF poderia dimerizar o TrkA e o p75NTR existe como um oligômero pré-formado que não é dissociado pelo NGF. Não há evidência para mostrar que TrkA e p75NTR interagem diretamente, então eles podem interagir indiretamente através da convergência de vias de sinalização a jusante e / ou compartilhar moléculas adaptadoras, ao invés de interações diretas receptor-receptor. Esses dados indicam que o destino final das células que coexpressam TrkA e p75NTR é complicado com a resposta funcional relacionada à abundância de cada receptor e dos diferentes agonistas. [9]
Receptores tipo Toll [TLRs]- Esta subclasse RTK é ativada por moléculas derivadas de patógenos, permitindo que o corpo detecte patógenos indesejados desde o início, bem como sinais de “perigo” de detecção que levam à eventual destruição desses patógenos. [2] Os TLRs sinalizam através do recrutamento de moléculas adaptadoras específicas, levando à ativação dos fatores de transcrição NF-κB e IRFs, que ditam o resultado das respostas imunes inatas. Os TLRs são expressos em células imunes inatas, como células dendríticas (DCs) e macrófagos, bem como em células não imunes, como células de fibroblastos e células epiteliais. Os TLRs são amplamente classificados em duas subfamílias com base em sua localização, TLRs de superfície celular e TLRs intracelulares. Os TLRs de superfície celular incluem TLR1, TLR2, TLR4, TLR5, TLR6 e TLR10, enquanto os TLRs intracelulares estão localizados no endossomo e incluem TLR3, TLR7, TLR8, TLR9, TLR11, TLR12 e TLR13. Os TLRs de superfície celular reconhecem principalmente os componentes da membrana microbiana, como lipídios, lipoproteínas e proteínas. O TLR4 reconhece o lipopolissacarídeo bacteriano (LPS). TLR2 juntamente com TLR1 ou TLR6 reconhece uma grande variedade de PAMPs incluindo lipoproteínas, peptidoglicanos, ácidos lipotecóicos, zimosan, manana e tGPI-mucina. TLR5 reconhece a flagelina bacteriana. TLR10 é pseudogene em camundongo devido a uma inserção de um códon de parada, mas TLR10 humano colabora com TLR2 para reconhecer ligantes de listeria. O TLR10 também pode detectar a infecção pelo vírus influenza A. Os TLRs intracelulares reconhecem ácidos nucléicos derivados de bactérias e vírus e também reconhecem ácidos nucléicos próprios em condições de doença, como autoimunidade. O TLR3 reconhece RNA de fita dupla viral (dsRNA), pequenos RNAs interferentes e auto-RNAs derivados de células danificadas. TLR7 é predominantemente expresso em plasmocitoides DCs (pDCs) e reconhece RNA de fita simples (ss) de vírus. Ele também reconhece RNA de bactérias estreptococos B em DCs convencionais (cDCs). O TLR8 humano responde ao RNA viral e bacteriano. A análise estrutural revelou que o TLR8 humano não estimulado existe como um dímero pré-formado e, embora o Z-loop entre LRR14 e LRR15 seja clivado, as metades N- e C-terminais permanecem associadas entre si e participam no reconhecimento do ligante e dimerização. A ligação do ligante induz a reorganização do dímero para trazer os dois terminais C em estreita proximidade. O TLR13 reconhece rRNA 23S bacteriano e componentes desconhecidos do vírus da estomatite vesicular. O TLR9 reconhece o DNA bacteriano e viral rico em motivos CpG-DNA não metilados; também reconhece a hemozoína, um subproduto cristalino insolúvel gerado pelo Plasmodium falciparum durante o processo de desintoxicação após a digestão da hemoglobina do hospedeiro. O TLR11 está localizado no endolisossomo e reconhece a flagelina ou um componente proteico desconhecido da Escherichia coli uropatogênica (UPEC), bem como uma molécula semelhante à profilina derivada do Toxoplasma gondii. TLR12 é predominantemente expresso em células mieloides e é altamente semelhante ao TLR11 e reconhece a profilina de T. gondii. TLR12 funciona como um homodímero ou um heterodímero com TLR11. [1]
Essa pequena classe é similar aos RTKs em estrutura, porém fosforila resíduos de serina e/ou treonina em vez de tirosina.[10] Fosforilam especificamente as cadeias laterais de hidroxila de resíduos de aminoácidos de serina ou treonina. Essas quinases podem ter um papel na proliferação celular, diferenciação, apoptose e até mesmo no desenvolvimento embrionário. Os ligantes proteicos, como o TFG-β, ativam uma família de receptores semelhantes às tirosinas-cinase receptoras, exceto por possuírem um domínio de cinase de serina/treonina na região citoplasmática da proteína. Existem duas isoformas da proteína receptora monomérica: tipo I (sete formas) e tipo II (cinco formas). No estado basal, essas proteínas são monômeros; depois do acoplamento de um ligante agonista, essas proteínas dimerizam e resultam na fosforilação do domínio de cinase do monômero tipo I, que ativa o receptor. Em seguida, o receptor ativado fosforila uma proteína reguladora gênica conhecida como Smad. Quando é fosforilada pelo receptor ativado de uma molécula de serina, a Smad dissocia-se do receptor, migra para o núcleo, combina-se com fatores de transcrição e regula os genes que resultam na morfogênese e transformação. Também existem Smads inibitórias (isoformas Smad6 e Smad7), que competem com as Smads fosforiladas para interromper a sinalização.[3]
As células expressam uma família de receptores de citocinas como γ-interferona e hormônios, como o hormônio do crescimento e a prolactina, que enviam sinais ao núcleo por uma via mais direta que as tirosinas-cinase receptoras. Esses receptores não têm atividade enzimática intrínseca; em vez disso, seu domínio intracelular liga-se a uma tirosina-cinase intracelular diferente conhecida como Jak. Com a dimerização induzida pelo acoplamento do ligante, as Jak fosforilam outras proteínas, conhecidas como STAT, que se transferem ao núcleo e regulam a transcrição. A via completa é conhecida como Jak-STAT (Gough et al., 2008; Wang et al., 2009). Nos mamíferos, existem 4 Jak e 6 STAT que, dependendo do tipo celular e do sinal, combinam-se diferentemente para ativar a transcrição dos genes. [3]