Winteraceae
Winteraceae é uma família de plantas com flor da ordem Canellales do grupo das magnoliídeas, que agrupa cerca de 93 espécies[5] de árvores e arbustos repartidas por 5 géneros.[3] As espécies desta famílias ocorrem em em regiões de clima tropical a temperado, principalmente no Hemisfério Sul, com distribuição natural numa vasta região que vai da Malésia, Oceânia, leste da Austrália até à Nova Zelândia, em Madagáscar e nos Neotrópicos,[6] com a maioria dos géneros concentrados na Australásia e na Malésia. A maioria das espécies ocorre em habitats de florestas húmidas, florestas alto-montanas ou em ambientes pantanosos.[7] Muitos dos membros da família são fragrantes e são utilizados na produção de óleos essenciais.
| Winteraceae | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Albiano - recente[1][2] | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Géneros[3][4] | |||||||||||||
| Sinónimos | |||||||||||||


.jpg)

.jpg)


_fruits_(Inao_V%C3%A1squez).jpg)

Descrição
editarMorfologia
editarOs membros da família Winteraceae são árvores ou arbustos lenhosos, com tecidos ricas em óleos aromáticos, com xilema que revela uma estrutura anatómica composta inteiramente de traqueídeos longitudinais, parênquima axial, raios e com grandes pontoações nas folhas e no ritidoma.[8]
Apresentam folhas alternas, com pontos verdes claros e um aroma perfumado. As estípulas estão ausentes.
As flores são pequenas, geralmente bissexuais, aparecendo maioritariamente em inflorescências do tipo cimeira ou fascículo. Têm duas a seis sépala livres e valvadas, embora estejam unidas em Drimys.[6] As pétalas ocorrem em duas ou mais séries. As flores apresentam numerosos estames e um a vários carpelos.[9] O pólen é monoaperturado.
Apresentam carpelos plicados, caracteres utilizados por muito tempo para considerar as Winteraceae como uma das famílias mais primitivas do grupo das angiospermas. Em todas as espécies desta família, os caracteres distintivos das tétrades de pólen libertadas são facilmente reconhecidos utilizando microscopia de luz e eletrónica.[10][11]
A família Winteraceae é caracterizada pela ausência de elementos de vaso no seu xilema,[12] tendo em seu lugar traqueídeos. Esta característica torna estas plantas relativamente imunes a embolias do xilema causadas por temperaturas negativas. Além disso, a oclusão vascular pode ocorrer perto das aberturas dos estomas, impedindo a entrada de água em excesso.[3] Há assim algumas evidências de que os traqueídeos oferecem algumas vantagens sobre os vasos do xilema, pois evitam o embolismo devido ao congelamento, o que permitiria retenção de folhas e de alguma fotossíntese em condições de inverno.
Os principais traços morfológicos da família Winteraceae são os seguintes:
- Pequenas árvores até 13 m de altura ou arbustos (por vezes não superior a 10 cm de altura), perenes, glabras ou por vezes com papilas piliformes ou tricomas minúsculos, aromáticas, raramente epífitas ou trepadoras (em Drimys).
- Folhas espiraladas, raramente subverticiladas, simples, inteiras, geralmente pecioladas, pinatinervadas, broquidódromas, sem estípulas, frequentemente com glândulas translúcidas minúsculas, face inferior frequentemente glaucosa, supervolutas. Estomas paracíticos.
- Caules com ramificação simpodial ou monopodial, botões sem escamas (exceto em Drimys). Vasoss do xilema ausentes. Nó bi- ou trilacunado, com 3 traços foliares. São comuns estolões e ramos ladrões. Casca aromática, muitas vezes com um sabor pungente ou mais raramente amargo.
- Inflorescências terminais, intercalares ou axilares, inflorescências parciais nas axilas das brácteas agrupadas formando dicásios simples ou compostos, frequentemente reduzidas a tríades, ou flores solitárias.
- Flores brancas a amarelas ou vermelhas, perfeitas (unissexuais em Tasmannia), actinomorfas a dissimétricas, hipóginas, com as diferentes peças em espiral ou aparentemente em espirais. Recetáculo curto ou longo. Disco hipogínico ausente. Cálice caliptrado, com uma pequena abertura apical fechada por 2-4(-6) lóbulos em verticilo, quebrando cedo e persistente (e.g. em Zygogynum) ou então quebrando na antese e decíduo (e.g. em Drimys). Pétalas (0-)2-50, em 3 verticilos, inteiras, livres ou as exteriores soldadas e rompendo na antese (Zygogynum), o seu número é variável mesmo na mesma planta. Androceu de 3-100 estames em 3-5 verticilos, livres, em número variável na mesma planta, filamentos curtos e grossos ou longos e cilíndricos, anteras 4 esporangiadas (2 esporangiadas em Zygogynum), teca apical e basifixa à lateral, deiscência lateroextrorsa por fenda longitudinal, conectivo pouco desenvolvido, apicalmente proeminente em Zygogynum. Gineceu suculento, com 1-50 carpelos em verticilo, apocárpico a variadamente sincarpado, estilo ausente, estigma 1 por carpelo, (sub)sésseis, circulares a lineares, adaxiais, por vezes estendendo-se ao longo de toda a margem adaxial do carpelo, ou 1 por ovário, capitado (Takhtajania), geralmente húmido e papiloso; óvulos (1-)2-muitos em geralmente 1 fileira em cada uma das 2 placentações parietais, lineares, opostas, paralelas ao estigma, anatrópicas, apotróficas, bitégmicas, crassinuceladas, com opérculo nuclear, ou placentações axiais em ovários sincarpados.
- Fruto em baga, por vezes muito duro devido aos esclereídeos, com ou sem polpa; em ovários sincarpados, por vezes fundidos num sincarpo; em Takhtajania o fruto é aparentemente uma cápsula septada.
- Sementes poucas a muitas, geralmente obovóides, de eixo reto ou curvo, lisas a pustulosas, com testa cinzento-escura a negra, duras, quebradiças, endosperma abundante, oleoso, embrião muito pequeno, ovoide e ligeiramente bilobado apicalmente.
- Pólen geralmente em tétrades, tetraédrico, raramente em mónadas (em alguns Zygogynum), citoplasmas separados por ectexina ou em contacto; grãos geralmente com um poro distal, raramente unicolpados (tricotomocolpados), ectexina bem desenvolvida distalmente, com um teto reticulado, columelas e camada basal, apenas esta última presente nos septos de separação de cada grão tétrade paredes rectas ou onduladas, columelas em uma ou raramente em duas filas, lumina grande e irregular, endexina e ectexina finas, gradualmente espessadas em direção à abertura, poros irregulares com um bordo espessado de endexina, intina por vezes saliente numa cúpula a partir da abertura.
- Número cromossómico: dois números básicos presentes: Zygogynum, Pseudowintera e Drimys têm n = 43, com tetraploidia ocasional e cromossomas pequenos, enquanto Tasmannia tem n = 13, com tetraploidia ocasional e cromossomas médios; Takhtajania tem n = 18.
Fitoquímica
editarOs membros de Winteraceae apresentam cristais de oxalato de cálcio no mesófilo das folhas. Óleos essenciais e resinas isoprenóides amplamente distribuídos pela planta, pelo que muitas têm um sabor picante. Raros alcalóides. Estão presentes flavonas, flavonóis e flavanonóis (como a dihidroquercetina), bem como taninos e proantocianidinas. Alguns representantes são acumuladores de alumínio.
O género Drimys é cianogenético.
Ocorrência
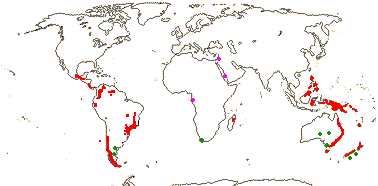
editarAs Winteraceae são uma família associada essencialmente ao hemisfério sul e à flora antárctica e podem ser encontradas geralmente em regiões húmidas temperadas e subtropicais do hemisfério sul e também a altitudes mais elevadas nos trópicos húmidos, desde regiões tropicais a regiões temperadas da Malésia, ecozona da Oceânia, leste da Austrália, Nova Zelândia, Madagáscar e da região neotropical. Esta família desapareceu do registo fóssil de África há aproximadamente 24 milhões de anos.
As plantas desta família ocorrem principalmente no hemisfério sul, e são encontradas em regiões de clima tropical a temperado da Malésia, Oceânia, leste da Austrália, Nova Zelândia, Madagáscar e nos Neotrópicos,[6] com a maioria dos géneros concentrados na Australásia e na Malésia, região onde se localiza o centro de diversidade do grupo.
Os cinco géneros, Takhtajania, Tasmannia, Drimys, Pseudowintera e Zygogynum s.l., têm todos populações geográficas distintas. Takhtajania inclui uma única espécie, Takhtajania perrieri, endémica em Madagáscar; Tasmannia tem a maior distribuição de géneros em Winteraceae, com espécies nas Filipinas, Bornéu, Nova Guiné, leste da Austrália e Tasmania; Drimys encontra-se no domínio neotropical, desde o sul do México até às florestas subárcticas do sul da América do Sul; Pseudowintera encontra-se apenas na Nova Zelândia; e Zygogynum tem espécies na Nova Guiné e na Nova Caledónia.[13]
Ecologia
editarAs flores das Winteraceae têm um odor moderado a forte, que varia de perfumado a frutado ou a peixe. As flores bissexuais são protândricas, com os estigmas a segregarem néctar na fase feminina enquanto os estames estão fechados, enquanto na fase masculina os filamentos estaminados aumentam de tamanho e as anteras abrem-se, enquanto os estigmas já não estão receptivos. As fases podem ou não sobrepor-se.
As flores geralmente fecham à noite, mas não em Pseudowintera ou em algumas Tasmannia. As flores podem durar de algumas horas a 12 dias. Num espécime o número de flores abertas pode variar de 2 a várias centenas.
Diferentes tipos de insectos visitam as flores, sendo particularmente interessante a relação de espécies de Sabatinca (Lepidoptera, Micropterygidae) com a polinização de Zygogynum baillonii e Zygogynum bicolor. Em alguns casos, a polinização é anemófila.
Algumas espécies são auto-compatíveis e a apomixia é conhecida em Drymis piperita. Alguns pássaros estão envolvidos na dispersão dos frutos em Pseudowintera e Drimys brasiliensis, mas em Zygogynum os frutos são libertados na árvore. A reprodução por estratificação e estolonização é por vezes importante em Drimys.
As espécies da família Winteraceae fazem parte do dossel florestal e do sub-bosque das florestas subtropicais e tropicais de baixa a média altitude, por vezes em florestas e charnecas temperadas montanas (até 4175 m de altitude) ou marítimas.
Filogenia e taxonomia
editarFilogenia
editarAnálises filogenéticas posicionaram a família Winteraceae na ordem Canellales como grupo-irmão de Canellaceae, família de espécies arbóreas que possuem elementos de vaso, sendo indicativo que a ausência pode ser um estado apomórfico e não plesiomórfico[14].
Esta família é de particular interesse filogenético por ser grupo de angiospérmicas tão primitivas, distantemente relacionada com as Magnoliaceae, embora tenha uma distribuição muito mais meridional.[6] Estima-se que esta família tenha existido desde há 105 e, pelo menos, desde há 35 milhões de anos atrás.[3][10]
Sendo uma das poucas angiospérmicas que formam tétrades persistentes com escultura proeminente, o pólen de Winteraceae é raro mas fácil de identificar no registo fóssil.[10] Amostras de pólen (Walkeripollis) encontradas em depósitos do Cretácio no Gabão podem indicar que a família tem pelo menos 120 milhões de anos,[15] mas a associação desses fósseis com Winteraceae é incerta.[10] Conhecem-se também fósseis de pólen desta família do Aptiano Superior-Baixo Albiano (há 125 Ma) e fósseis de madeira (Winteroxylon) do Santoniano-Campaniano (85-70 Ma) da ilha James Ross, na Antárctida.
Se conoce polen fósil de esta familia del Cretácico de Gabón y del Aptiense superior-Albiense inferior (hace 125 Ma) y madera fósil (Winteroxylon) del Santoniense-Campaniense (85-70 Ma) de la isla James Ross en la Antártida.
Os fósseis mais antigos e inequívocos de Winteraceae são do meio para o final do Albiano de Israel (~110 milhões de anos; descritos como Qatanipollis). [2] Fósseis de pólen indicam que a distribuição foi muito mais ampla do que é agora,[3] atingindo o norte até a Gronelândia durante o Paleoceno (Daniano),[10] e desaparecendo da África continental (Península do Cabo, África do Sul) no Mioceno.[16]
Igualmente caraterística é a madeira de Winteraceae, que carece de vasos de xilema, em contraste com a maioria das outras plantas com flor.[12] A madeira fóssil de Winteraceae foi encontrada em depósitos datados do Cretáceo Superior ao Paleogénico (c. 85-35 milhões de anos atrás) da Antártida (Santoniano-Campaniano),[17] oeste da América do Norte (Central Valley, Califórnia; Maastrichtiano)[18] e Europa (Helmstedt, Alemanha; Eoceno).[19]
Posição filogenética da família
editarAs Winteraceae foram inicialmente colocadas como um grupo basal dentro das angiospérmicas devido à sua madeira sem vasos.[20] Os vasos do xilema foram vistos como um importante caráter evoluído para a diversificação e sucesso das angiospermas, de modo que a madeira sem vasos foi vista como uma caraterística arcaica, resultando na colocação basal das Winteraceae. No entanto, o progresso da filogenia molecular colocou as Winteraceae dentro das magnoliídeas, bem dentro das angiospermas.[20] Esta colocação sugere que a madeira sem vasos das Winteraceae foi um carácter derivado e não ancestral.
Através do registo de pólen fóssil, é possível postular que as Winteraceae se tenham movido do norte de Gondwana através do sul de Gondwana no Cretáceo.[21] Isto significou o movimento de ambientes húmidos quentes para ambientes húmidos temperados onde ocorreram eventos de congelamento e descongelamento. A madeira sem vasos tem 20% da condutividade da água da madeira com vasos, no entanto, sob eventos de congelamento, a madeira com vasos perde até 85% da condutividade da água, enquanto a madeira sem vasos perde no máximo 6% da condutividade de água.[22]
A capacidade de evitar uma limitação hídrica grave e, por conseguinte, a queda de folhas, é hipotetizada como uma pressão evolutiva importante por trás da reversão para a madeira sem vasos. Este facto é ainda apoiado pela hipótese da heteroxila, em que os vasos "primitivos" conferiam pouca diferença na eficiência hidráulica do caule em condições normais em comparação com as angiospérmicas sem vasos. Isto indicaria que a pressão dos eventos de congelamento e descongelamento, e o subsequente risco de embolia, seriam um fator evolutivo mais forte em comparação com as restrições hidráulicas mais fracas da madeira sem vasos em comparação com os vasos "primitivos".[23]
Este movimento de ambientes quentes e húmidos para ambientes húmidos com temperaturas onde ocorreram eventos de congelamento e descongelamento é visto como a pressão evolutiva por trás da reversão única para madeira sem vasos que ocorreu em Winteraceae.
Outra caraterística das Winteraceae que foi considerada como indicando uma posição basal na filogenia foi a presença de tampões estomáticos cerosos, considerados como limitando a perda de água na respiração e, portanto, uma caraterística arcaica para limitar a perda de água.[24]
No entanto, estudos posteriores mostraram que, nestes ambientes húmidos, a cobertura de água na superfície das folhas diminui as taxas fotossintéticas e que os tampões estomáticos cerosos reduzem esta cobertura de água, reduzindo assim os impactos negativos na capacidade fotossintética.[25] Espécimes de Winteraceae com tampões estomáticos removidos viram diminuições nas taxas fotossintéticas de até 40%.[21] Isto mostra ainda que caracteres outrora considerados arcaicos podem ser adaptações derivadas a ambientes húmidos temperados.
Classificação
editarAs Winteraceae são um grupo primitivo de angiospermas, que, dependendo do sistema de classificação e dos caracteres considerados, foram incluídas no grupo Polycarpicae ou foram colocadas perto das Schisandraceae ou das Canellaceae.[26]
De acordo com o sistema APG I, de 1998, a família foi mantida em incertae sedis, não sendo atribuída a qualquer ordem,[27] mas desde então foi colocado em Canellales pelo sistema APG II, ordem onde foi mantida pelo sistema APG III e pelo sistema APG IV.[28][29][30]
Na sua presente circunscrição taxonómica, a família Winteraceae encontra-se subdividida em duas subfamílias (Takhtajanoideae (monotípica) e Winteroideae), com os seguintes géneros:[31]
- Takhtajanoideae
- Winteroideae
- Drimys J.R.Forster & G.Forster — sin.: Wintera Murray; Winterania P. & K; Winterana Medikus.
- Pseudowintera Dandy — sin.: Wintera van Tieghem.
- Tasmannia R.Brown
- Zygogynum Baillon — sin.: Belliolum van Tieghem; Bubbia van Tieghem; Exospermum van Tieghem; Tetrathalamus Lauterb..
Na circunscrição atrás apontada, a família Winteraceae apresenta a estrutura filogenética que consta do seguinte cladograma:
| Winteraceae |
| ||||||||||||||||||||||||
A família Winteraceae foi estabelecida em 1830 por Robert Brown ex John Lindley: An Introduction to the Natural System of Botany. 26 pp.[32] O género homónimo da família, Wintera, está incluído em Drimys. O nome do género Wintera Murray é um nome ilegítimo para Drimys. J.R.Forst. & G.Forst.[33]
A família Winteraceae R.Br. ex Lindl. nom. cons. divide-se em duas subfamílias: Taktajanioideae Leroy (sin.: Takhtajaniaceae J.-F. Leroy); e Winteroideae Arnott (sin.: Wintereae Meisn., Drimyidoideae Raf., Drimyideae Baill., Exospermeae Hutch., Temoideae Raf.).[34]
As Winteraceae desenvolvem-se em regiões temperadas a tropicais, especialmente nas regiões montanhosas dos trópicos. Encontram-se desde a Malásia até ao região do Pacífico, no leste da Austrália e Nova Zelândia, mas também na América Central e América do Sul, bem como em Madagáscar.
Existem quatro a sete géneros na família Winteraceae, dependendo da fonte. De acordo com Kew Gardens[35] existem os seguintes cinco géneros:[36]
- Bubbia Tiegh.: Contém apenas uma espécie, mas também está incluída em Zygogynum:
- Bubbia howeana (F.Muell.) Tiegh.: Encontra-se apenas na Ilha de Lord Howe, a leste da Austrália.
- Drimys J.R.Forst. & G.Forst. (sin.: Wintera Murray): Dependendo do autor, são incluídas 9 a 21 espécies. As cerca de 5 a 14 espécies asiáticas são frequentemente separadas como um género separado Tasmannia R.Br. ex DC. das outras espécies sul-americanas Drimys. A Drimys pertence:
- Drimys andina (Reiche) R.A.Rodr. & Quezada: Encontra-se na Argentina e no Chile.[33]
- Drimys granadensis L.f.: Ocorre no sul do México, Guatemala, Honduras, Costa Rica, Panamá, Colômbia e Peru.[33]
- Drimys winteri J.R.Forst. & G.Forst.: É muito comum no Chile e na Argentina.[33]
- Exospermum Tiegh.: Contém apenas uma espécie:
- Exospermum stipitatum (Baill.) Tiegh.: Encontra-se na Nova Caledónia.
- Pseudowintera Dandy: As três a quatro espécies encontram-se na Nova Zelândia.
- Pseudowintera colorata (Raoul) Dandy: Do norte da Nova Zelândia.
- Takhtajania Baranova & J.-F. Leroy: colocada na sua própria subfamília Takhtajanioideae J.-F.Leroy ou mesmo na sua própria família Takhtajaniaceae (J.-F.Leroy) J.-F.Leroy. Contém apenas uma espécie:[34]
- Takhtajania perrieri (Capuron) Baranova & J.-F. Leroy: Nativa de Madagáscar.[34]
- Tasmannia R.Br. ex DC.: As cerca de três espécies ocorrem na Austrália. São também classificadas por alguns autores como Drimys. Entre elas:
- Tasmannia lanceolata (Poir.) A.C.Sm.
- Zygogynum Baill.: As cerca de seis espécies distribuem-se da Nova Guiné à Nova Caledónia.
Redescoberta do género Takhtajania
editarA espécie Takhtajania perrieri foi recolhida pela primeira vez em 1909 no Maciço de Manongarivo, no centro de Madagáscar, a uma altitude de 1700 metros. Em 1963, o botânico francês René Paul Raymond Capuron examinou a amostra de planta não identificada, que identificou como uma nova espécie, a que deu o nome de Bubbia perrieri, em homenagem ao botânico francês Henri Perrier de la Bâthie, classificando-a no género australásico Bubbia.
Em 1978, os botânicos Margarita Alexandrovna Baranova e Jean-François Leroy reclassificaram a planta no seu próprio género, Takhtajania, em homenagem ao botânico soviético Armen Takhtajan. Muitas expedições subsequentes para encontrar a espécie foram inúteis, mas em 1994 o colecionador de plantas malgaxe Fanja Rasoavimbahoaka recolheu um espécime na Reserva Especial de Anjahanaribe-Sud, a 150 km do local onde o espécime de 1909 fora recolhido, que George E. Schatz identificou em maio de 1997 como Takhtajania. Uma expedição posterior descobriu um grande bosque da espécie no local onde foi recolhido o segundo exemplar.[37]
Usos
editarA espécie Drimys winteri (a casca-de-anta, comercializada no mercado internacional como Winter's bark) é uma árvore esguia, nativa das florestas subpolares magelânicas e das florestas temperadas chuvosas valdivianas do Chile e Argentina. É cultivada como planta de jardim devido à sua casca cor de mogno-vermelho, pelas suas folhas verde brilhantes e conjuntos de flores branco e creme e de cheiro ajasminado. A casca foi historicamente utilizada para prevenir o escorbuto.[38]
A Tasmannia lanceolata, conhecida também como pimenta-da-tasmânia, é também cultivada como um arbusto ornamental e cada vez mais para servir de condimento. Tasmannia piperita é notável pela grande variedade de números de pétalas, estames e pistilos, sendo usada como planta ornamental.
A espécie mais conhecida desta família é o canelo, Drimys winteri, nativo do Chile e da Argentina, cuja casca é utilizada como especiaria e como medicamento, pelas suas qualidades anti-escorbúticas, entre outras, e que é também o foiye, a planta sagrada dos Mapuche. É também utilizada na decoração e na construção de instrumentos musicais.
Na Nova Zelândia, a madeira da Pseudowintera axillaris é utilizada para interiores. Na Austrália, os frutos da Tasmannia purpurascens são utilizados como condimento pelo seu sabor picante, assim como os frutos e as folhas da Tasmannia lanceolata.
La especie más conocida de esta familia es el canelo, Drimys winteri, originario de Chile y Argentina, cuya corteza se usa como especia y como fármaco, por sus cualidades antiescorbúticas, entre otras, y que es además el foiye, la planta sagrada de los mapuches. También se usa en decoración y en la construcción de instrumentos musicales. En Nueva Zelanda se usa la madera de Pseudowintera axillaris para interiores. En Australia, los frutos de Tasmannia purpurascens se usan como condimento por su sabor picante, así como los frutos y las hojas de Tasmannia lanceolata.
Ocorrência no Brasil
editarNo Brasil é representada por apenas um gênero (Drimys), distribuído pela Caatinga, Cerrado e Mata Atlântica, não sendo uma espécie endêmica. Para grande parte dos autores sete espécies ocorreriam no Brasil, sendo elas a D. brasiliensis, D. angustifolia, D. granadensis, D. montana, D. retorta, D. roraimensis e D. winteri. Com base em observações de campo, pôde-se verificar que no Rio Grande do Sul, enquanto D. angustifolia floresce de julho a novembro com pico de floração em setembro, D. brasiliensis apresenta uma floração praticamente contínua ao longo do ano, com dois picos: janeiro-fevereiro e julho-setembro.
Marquínez (2009) e BFG (2015) reconhecem para o Brasil três espécies: D. brasiliensis com três subespécies [D. brasiliensis subsp. brasiliensis, D. brasiliensis subsp. subalpina Ehrend. & Gottsb. e D. brasiliensis subsp. sylvatica (A.St.-Hil.) Ehrend. & Gottsb.], D. angustifolia e D. roraimensis, em concordância com Ehrendorfer et al. (1979). As espécies brasileiras estão distribuídas geograficamente nas seguintes Regiões e Estados: Norte (Roraima), Nordeste (Bahia), Centro-Oeste (Distrito Federal) Sudeste (Espírito Santo, Minas Gerais, Rio de Janeiro e São Paulo) e no Sul (Paraná, Rio Grande do Sul e Santa Catarina)[39].
Espécies brasileiras
editar
No Brasil ocorrem as seguintes espécies:
- Drimys angustifolia Miers
- Drimys brasiliensis Miers
- Drimys brasiliensis Miers subsp. brasiliensis
- Drimys brasiliensis subsp. subalpina Ehrend. & Gottsb.
- Drimys brasiliensis subsp. sylvatica (A.St.-Hil.) Ehrend. & Gottsb.
- Drimys brasiliensis var. campestris (A.St.-Hil.) Miers
- Drimys brasiliensis var. montana (Miers) Hauman
- Drimys brasiliensis var. ptaritepuiensis Steyerm.
- Drimys brasiliensis var. retorta (Miers) A.C.Sm.
- Drimys brasiliensis var. roraimensis A.C.Sm.
- Drimys brasiliensis var. sylvatica (A.St.-Hil.) Miers
- Drimys granadensis L.f.
- Drimys granadensis var. axillaris A.St.-Hil.
- Drimys granadensis var. campestris A.St.-Hil.
- Drimys granadensis var. montana A.St.-Hil.
- Drimys granadensis var. sylvatica A.St.-Hil.
- Drimys montana (A.St.-Hil.) Miers
- Drimys retorta Miers
- Drimys roraimensis (A.C.Sm.) Ehrend. & Gottsb.
- Drimys winteri J.R.Forst. & G.Forst.
- Drimys winteri var. semiglobosa Dusén
Referências
editar- ↑ «Canellales». www.mobot.org. Consultado em 20 de julho de 2023
- ↑ a b WALKER, J. W.; BRENNER, G. J.; WALKER, A. G. (17 de junho de 1983). «Winteraceous Pollen in the Lower Cretaceous of Israel: Early Evidence of a Magnolialean Angiosperm Family». Science. 220 (4603): 1273–1275. Bibcode:1983Sci...220.1273W. ISSN 0036-8075. PMID 17769368. doi:10.1126/science.220.4603.1273
- ↑ a b c d e Stevens, P.F. «Winteraceae». Angiosperm Phylogeny Website
- ↑ Winteraceae em APWeb
- ↑ «Winteraceae R.Br. ex Lindl. | Plants of the World Online | Kew Science». Plants of the World Online. Consultado em 26 de novembro de 2021
- ↑ a b c d Hutchinson (1973). The Families of Flowering Plants. [S.l.]: Oxford at the Clarendon Press
- ↑ Judd, W.S.; Campbell, C.S.; Kellogg, E.A. & Stevens, P.F. 1999. Plant systematics: A Phylogenetic Approach. Sinauer Associates, Sunderland, Massachussets. 464p.
- ↑ DE ABREU, DANIELA CLEIDE AZEVEDO et al. Caracterização morfológica de frutos e sementes de cataia (Drimys brasiliensis Miers.-Winteraceae). Embrapa Florestas-Artigo em periódico indexado (ALICE), 2005.
- ↑ WINTERACEAE. [S. l.], 2010. Disponível em: https://www.britannica.com/plant/Winteraceae. Acesso em: 13 nov. 2019.
- ↑ a b c d e Grímsson, Friðgeir; Grimm, Guido W.; Potts, Alastair J.; Zetter, Reinhard; Renner, Susanne S. (2018). «A Winteraceae pollen tetrad from the early Paleocene of western Greenland, and the fossil record of Winteraceae in Laurasia and Gondwana». Journal of Biogeography (em inglês). 45 (3): 567–581. ISSN 1365-2699. doi:10.1111/jbi.13154
- ↑ an der Ham, Raymond; Joan van Heuven, Bertie (2002). «Evolutionary trends in Winteraceae pollen». Grana. 41 (1): 4–9. doi:10.1080/00173130260045431
- ↑ a b Taylor S. Feild, Tim Brodribb & N. Michele Holbrook (2002). «Hardly a relict: freezing and the evolution of vesselless wood in Winteraceae» (PDF). Evolution. 56 (3): 464–478. PMID 11989678. doi:10.1554/0014-3820(2002)056[0464:HARFAT]2.0.CO;2
- ↑ Marquínez, Xavier; Lohmann, Lúcia G.; Salatino, Maria L. Faria; Salatino, Antonio; González, Favio (Novembro 2009). «Generic relationships and dating of lineages in Winteraceae based on nuclear (ITS) and plastid (rpS16 and psbA-trnH) sequence data». Molecular Phylogenetics and Evolution (em inglês). 53 (2): 435–449. PMID 19580880. doi:10.1016/j.ympev.2009.07.001
- ↑ HERTZOG, Anelise et al. Winteraceae do Rio Grande do Sul, Brasil. Rodriguésia, [S. l.], p. 1-10, 9 nov. 2015. DOI 10.1590/2175-7860201667113. Disponível em: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S2175-78602016000100251. Acesso em: 9 nov. 2019.
- ↑ Doyle, J. A. 1999. The rise of angiosperms as seen in the African Cretaceous record. Pp. 3-29, in Scott, L., Cadman, A., & Verhoeven, R. (eds), Proceedings of the Third Conference on African Palynology, Johannesburg, 14–19 September 1997. A. A. Balkema, Rotterdam.
- ↑ Coetzee, J. A.; Praglowski, J. (1988). «Winteraceae pollen from the miocene of the southwestern cape (south africa)». Grana. 27 (1): 27–37. ISSN 0017-3134. doi:10.1080/00173138809427730

- ↑ Poole, I (2000). «The First Record of Fossil Wood of Winteraceae from the Upper Cretaceous of Antarctica». Annals of Botany. 85 (3): 307–315. ISSN 0305-7364. doi:10.1006/anbo.1999.1049
- ↑ Page, Virginia M. (1979). «Dicotyledonous wood from the Upper Cretaceous of central California». Journal of the Arnold Arboretum. 60 (3): 223–249. doi:10.5962/p.185886
- ↑ Gottwald, H (1992). «Hölzer aus marinen Sanden des oberen Eozän von Helmstedt (Niedersachsen)». Palaeontographica Abteilung B. 225: 27–103
- ↑ a b Soltis, Pamela S.; Soltis, Douglas E. (outubro 2004). «The origin and diversification of angiosperms». American Journal of Botany (em inglês). 91 (10): 1614–1626. ISSN 0002-9122. PMID 21652312. doi:10.3732/ajb.91.10.1614
- ↑ a b Feild, Taylor S.; Zwieniecki, Maciej A.; Holbrook, N. M. (2000). «Winteraceae Evolution: An Ecophysiological Perspective». Annals of the Missouri Botanical Garden. 87 (3). 323 páginas. JSTOR 2666192. doi:10.2307/2666192
- ↑ Feild, Taylor S.; Brodribb, Tim; Holbrook, N. Michele (2002). «Hardly a Relict: Freezing and the Evolution of Vesselless Wood in Winteraceae». Evolution. 56 (3): 464–478. ISSN 0014-3820. JSTOR 3061586. PMID 11989678. doi:10.1111/j.0014-3820.2002.tb01359.x
- ↑ Hudson, Patrick J.; Razanatsoa, Jacqueline; Feild, Taylor S. (janeiro 2010). «Early vessel evolution and the diverisification of wood function: Insights from Malagasy Canellales». American Journal of Botany (em inglês). 97 (1): 80–93. ISSN 0002-9122. PMID 21622369. doi:10.3732/ajb.0900130
- ↑ Cronquist, Arthur (1981). An integrated system of classification of flowering plants. [S.l.: s.n.] ISBN 0-231-03880-1. OCLC 180644501
- ↑ Liang, Xiao-Qing; Lu, Ping; Zhang, Jian-Wei; Su, Tao; Zhou, Zhe-Kun (setembro 2018). «First fossils of Zygogynum from the Middle Miocene of Central Yunnan, Southwest China, and their palaeobiogeographic significance». Palaeoworld (em inglês). 27 (3): 399–409. doi:10.1016/j.palwor.2018.05.003
- ↑ AP-website.
- ↑ Angiosperm Phylogeny Group (1998). «An ordinal classification for the families of flowering plants». Annals of the Missouri Botanical Garden. 85 (4): 531–553. JSTOR 2992015. doi:10.2307/2992015
- ↑ Angiosperm Phylogeny Group (2003). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II». Botanical Journal of the Linnean Society. 141 (4): 399–436. doi:10.1046/j.1095-8339.2003.t01-1-00158.x
- ↑ Angiosperm Phylogeny Group (2009). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III». Botanical Journal of the Linnean Society. 161 (2): 105–121. ISSN 0024-4074. doi:10.1111/j.1095-8339.2009.00996.x
- ↑ Angiosperm Phylogeny Group (2016). «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV». Botanical Journal of the Linnean Society. 181 (1): 1–20. ISSN 0024-4074. doi:10.1111/boj.12385
- ↑ APWeb: Winteraceae.
- ↑ «Winteraceae». Tropicos. Missouri Botanical Garden. 42000000
- ↑ a b c d «Winteraceae». Agricultural Research Service (ARS), United States Department of Agriculture (USDA). Germplasm Resources Information Network (GRIN)
- ↑ a b c Winteraceae auf der APWebsite.
- ↑ Datenblatt bei Plants of the World online des Board of Trustees of the Royal Botanic Gardens, Kew. Zuletzt eingesehen am 1. März 2020.
- ↑ J. L. R. Every, 2009: Neotropical Winteraceae. In: W. Milliken, B. Klitgĺrd, A. Baracat: Neotropikey - Interactive key and information resources for flowering plants of the Neotropics. Datenblatt.
- ↑ Schatz, George E. (2000). «The Rediscovery of a Malagasy Endemic: Takhtajania perrieri (Winteraceae)». Annals of the Missouri Botanical Garden. 87 (3): 297–302. JSTOR 2666189. doi:10.2307/2666189
- ↑ «Winteraceae: Plant family». Encyclopædia Britannica. Consultado em 26 de setembro de 2016
- ↑ Winteraceae in Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro. Disponível em: <http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB251>. Acesso em: 09 Nov. 2019
Bibliografia
editar- Grímsson, F., G. W. Grimm, A. Potts, R. Zetter, and S. S. Renner. (2017). A Winteraceae pollen tetrad from the early Paleocene of western Greenland, and the fossil record of Winteraceae in Laurasia and Gondwana. Journal of Biogeography 2017: 1–15.
- Hutchinson, J. (1964). The genera of flowering plants (Angiospermae) based principally on the Genera Plantarum of G. Bentham and J.D. Hooker. Vol. 1. [S.l.]: Oxford: Clarendon Press. 516 págs.
- Vink, W. (1993). «Winteraceae.». Kubitzki, K., Rohwer, J.G. & Bittrich, V. (Editores). The Families and Genera of Vascular Plants. II. Flowering Plants - Dicotyledons. [S.l.]: Springer-Verlag: Berlín. ISBN 3-540-55509-9
- Karol, K.G., Suh, Y., Schatz, G.E. & Zimmer, E.A. (2000). «Molecular evidence for the phylogenetic position of Takhtajania in the Winteraceae: inference from nuclear ribosomal and chloroplast gene spacer sequences». Annals of the Missouri Botanical Garden. 87 (3). p. 414-432
- Keating, R.C. (2000). «Anatomy of the young vegetative shoot of Takhtajania perrieri (Winteraceae)». Annals of the Missouri Botanical Garden. 87 (3). p. 335-346
- Watson, L., and Dallwitz, M.J. (1992). «The families of flowering plants: descriptions, illustrations, identification, and information retrieval. Version: 29th July 2006.». Consultado em 29 de novembro de 2006. Arquivado do original em 3 de janeiro de 2007
Ligações externas
editar- Chilean Winteraceae Chileflora
- Winteraceae em APWebsite.
- Winteraceae in L. Watson and M. J. Dallwitz (1992 onwards): The families of flowering plants.
- Mapa
- Frutos de Drimys winteri
- Winteraceae de Chile Chileflora
- Flor de Zygogynum baillonii
- Rama en flor de Takhtajania perrieri
- Rama en flor de Pseudowintera colorata
- Inflorescencia masculina de Tasmannia purpurascens
- Frutos y hojas de Tasmannia lanceolata
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}