Tyrannosauridae
Tyrannosauridae (ou tiranossaurídeos)[2] é uma família de dinossauros terópodes tetanúreos, característicos do Cretáceo superior. O grupo inclui os maiores predadores carnívoros que já existiram sobre a Terra, entre eles o famoso Tyrannosaurus rex. Seus fósseis foram encontrados em diversas regiões da América do Norte e da Ásia.
| Tyrannosauridae | |
|---|---|

| |
| Sentido horário a partir do canto superior esquerdo: Tyrannosaurus, Daspletosaurus, Tarbosaurus, Gorgosaurus, Zhuchengtyrannus e Alioramus | |
| Classificação científica | |
| Domínio: | Eukaryota |
| Reino: | Animalia |
| Filo: | Chordata |
| Clado: | Dinosauria |
| Clado: | Saurischia |
| Clado: | Theropoda |
| Superfamília: | †Tyrannosauroidea |
| Clado: | †Pantyrannosauria |
| Clado: | †Eutyrannosauria |
| Família: | †Tyrannosauridae Osborn, 1906 |
| Espécie-tipo | |
| †Tyrannosaurus rex Osborn, 1905
| |
| Subgrupos[1] | |
| Sinónimos | |
Uma característica distintiva do grupo são os membros anteriores, minúsculos por relação ao resto do corpo e demasiado pequenos para desempenharem uma função activa. Os tiranossaurídeos mediam, conforme o tipo, entre 8 a 15 metros de comprimento e pesavam entre, dos menores, 2 toneladas[3] até os maiores, que de acordo com as mais modernas estimativas, pesavam entre cerca de 8,4 tonelada métricas (9,3 tonelada curtas) a 14 tonelada métricas (15,4 tonelada curtas).[3][4][5]
Ao contrário da maioria dos outros grupos de dinossauros, restos muito completos foram descobertos para a maioria dos tiranossaurídeos conhecidos. Isso permitiu uma variedade de pesquisas em sua biologia. Os estudos científicos têm se concentrado em sua ontogenia, biomecânica e ecologia, entre outros assuntos.
História das descobertas
editar
Os primeiros restos de tiranossaurídeos foram descobertos durante expedições lideradas pelo Serviço Geológico do Canadá, que localizou vários dentes espalhados. Esses dentes distintos de dinossauros receberam o nome de Deinodon ("dente terrível") por Joseph Leidy em 1856. Os primeiros espécimes bons de um tiranossaurídeo foram encontrados na Formação Horseshoe Canyon de Alberta, Canadá e consistiam em crânios quase completos com esqueletos parciais. Esses restos foram estudados pela primeira vez por Edward Drinker Cope em 1876, que os considerou uma espécie do tiranossauróide oriental Dryptosaurus. Em 1905, Henry Fairfield Osborn reconheceu que os restos de Alberta diferiam consideravelmente do Dryptosaurus e cunhou um novo nome para eles: Albertosaurus sarcophagus ("lagarto comedor de carne de Alberta").[6] Cope descreveu mais material de um tiranossaurídeo em 1892, na forma de vértebras isoladas, e deu a esse animal o nome de Manospondylus gigas. Essa descoberta foi negligenciada por mais de um século e causou polêmica no início dos anos 2000, quando foi descoberto que este material realmente pertencia e tinha prioridade de nome sobre o Tyrannosaurus rex.[7]
Em seu artigo de 1905 intitulado Albertosaurus, Osborn descreveu dois espécimes adicionais de tiranossaurídeos que foram coletados em Montana e Wyoming durante uma expedição de 1902 ao Museu Americano de História Natural, liderada por Barnum Brown. Inicialmente, Osborn considerou essas espécies como espécies distintas. O primeiro, ele chamou Dynamosaurus imperiosus ("lagarto do poder do imperador"), e o segundo, Tyrannosaurus rex ("lagarto rei tirano"). Um ano depois, Osborn reconheceu que esses dois espécimes, na verdade, vinham da mesma espécie. Apesar de o Dynamosaurus ter sido encontrado primeiro, o nome Tyrannosaurus apareceu uma página antes em seu artigo original descrevendo os dois espécimes. Portanto, de acordo com o Código Internacional de Nomenclatura Zoológica (ICZN), o nome Tyrannosaurus foi usado.[8]
Barnum Brown passou a coletar vários outros espécimes de tiranossaurídeos de Alberta, incluindo o primeiro a preservar os membros anteriores encurtados de dois dedos característicos do grupo (que Lawrence Lambe chamou de Gorgosaurus libratus, "lagarto feroz equilibrado", em 1914). Uma segunda descoberta significativa atribuída ao Gorgosaurus foi feita em 1942, na forma de um crânio completo bem preservado, embora excepcionalmente pequeno. O espécime esperou até o final da Segunda Guerra Mundial para ser estudado por Charles W. Gilmore, que o batizou de Gorgosaurus lancenis.[6] Este crânio foi re-estudado por Robert T. Bakker, Phil Currie e Michael Williams em 1988 e atribuído ao novo gênero Nanotyrannus.[9] Foi também em 1946 que paleontólogos da União Soviética iniciaram expedições à Mongólia e descobriram os primeiros restos de tiranossaurídeos da Ásia. Evgeny Maleev descreveu novas espécies mongóis de Tyrannosaurus e Gorgosaurus em 1955, e um novo gênero: Tarbosaurus ("lagarto assustador"). Estudos subsequentes, entretanto, mostraram que todas as espécies de tiranossaurídeos de Maleev eram na verdade uma espécie de Tarbosaurus em diferentes estágios de crescimento. Uma segunda espécie de tiranossaurídeo mongol foi encontrada mais tarde, descrita por Sergei Kurzanov em 1976, e recebeu o nome de Alioramus remotus ("ramo diferente remoto"), embora seu status como um verdadeiro tiranossaurídeo e não um tiranossauróide mais primitivo ainda seja controverso.[6][10]
Descrição
editar
Os tiranossaurídeos eram todos animais grandes, com todas as espécies capazes de pesar pelo menos 1 tonelada.[11] Um único espécime de Alioramus de um indivíduo estimado entre 5 e 6 metros de comprimento foi descoberto,[10] embora seja considerado por alguns especialistas como um jovem.[11][12] Albertosaurus, Gorgosaurus e Daspletosaurus mediam entre 8 e 10 metros de comprimento,[13] enquanto o Tarbosaurus alcançava comprimentos de 12 metros do focinho à cauda.[14] O enorme Tyrannosaurus rex atingiu 12,3 metros em um dos maiores espécimes, FMNH PR2081.[3]

Uma característica marcante da família é o crânio relativamente grande para seu tamanho.[15] Mediam entre 15 centímetros e 1,5 metro e abrigavam fortíssimos músculos, gerando uma mordida muito potente. Suas órbitas oculares eram grandes, o que indica uma boa visão. Seu olfato é provavelmente o mais apurado dentre todos os seres vivos que já existiram.[16] Os crânios tiranossaurídeos tinham muitas características únicas, incluindo ossos parietais fundidos com uma crista sagital proeminente, que corria longitudinalmente ao longo da sutura sagital e separava as duas fenestras supratemporais no teto do crânio. Atrás dessas fenestras, os tiranossaurídeos tinham uma crista nucal caracteristicamente alta, que também surgia dos parietais, mas corria ao longo de um plano transversal em vez de longitudinalmente. A crista nucal foi especialmente bem desenvolvida nos Tyrannosaurus, Tarbosaurus e Alioramus. Albertosaurus, Daspletosaurus e Gorgosaurus tinham cristas altas na frente dos olhos nos ossos lacrimais, enquanto Tarbosaurus e Tyrannosaurus tinham ossos pós-orbitais extremamente espessos formando cristas em forma de meia-lua atrás dos olhos. Alioramus tinha uma fileira de seis cristas ósseas no topo de seu focinho, surgindo dos ossos nasais; cristas inferiores foram relatadas em alguns espécimes de Daspletosaurus e Tarbosaurus, bem como no Appalachiosaurus, um tiranossauroide mais basal.[12][15]
Dentição
editar

Os tiranossaurídeos, como seus ancestrais tiranossauróides, eram heterodontes, com dentes pré-maxilares em forma de D em seção transversal e menores do que o resto. Ao contrário dos tiranossauróides anteriores e da maioria dos outros terópodes, os dentes maxilares e mandibulares de tiranossaurídeos maduros não são semelhantes a lâminas, mas extremamente espessos e frequentemente circulares em seção transversal, com algumas espécies tendo serrilhas reduzidas.[11] A contagem de dentes tende a ser consistente dentro das espécies, e as espécies maiores tendem a ter contagens de dentes mais baixas do que as menores. Por exemplo, Alioramus tinha 76 a 78 dentes em suas mandíbulas, enquanto o Tyrannosaurus tinha entre 54 e 60.[17]
William Abler observou em 2001 que as serrilhas dos dentes do Albertosaurus se assemelham a uma rachadura no dente terminando em um vazio redondo chamado ampola.[18] Os dentes de tiranossaurídeos eram usados para arrancar a carne de um corpo, então quando um destes puxasse um pedaço de carne, a tensão poderia causar uma serragem puramente parecida com uma rachadura se espalhar pelo dente.[18] No entanto, a presença da ampola teria distribuído essas forças por uma área de superfície maior e diminuído o risco de danos ao dente sob tensão.[18] A presença de incisões terminando em vazios tem paralelos na engenharia humana. Os fabricantes de guitarras usam incisões que terminam em vazios para, como Abler descreve, "conferir regiões alternadas de flexibilidade e rigidez" à madeira com que trabalham.[18] O uso de uma broca para criar uma espécie de "ampola" e evitar a propagação de rachaduras através do material também é usado para proteger as superfícies do avião.[18] Abler demonstrou que uma barra de acrílico com incisões chamadas "kerfs" e orifícios perfurados era mais de 25% mais forte do que uma com apenas incisões colocadas regularmente.[18] Ao contrário dos tiranossaurídeos e outros terópodes, os predadores antigos como os phytossauros e o Dimetrodon não tinham adaptações para evitar que as serrilhas parecidas com rachaduras de seus dentes se espalhassem quando submetidos às forças da alimentação.[18]
Esqueleto pós-craniano
editar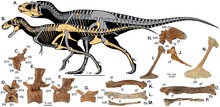
O crânio estava empoleirado na extremidade de um pescoço grosso em forma de S, e uma cauda longa e pesada atuava como um contrapeso para equilibrar a cabeça e o tronco, com o centro de massa sobre os quadris. Os tiranossaurídeos são conhecidos por seus membros anteriores de dois dedos proporcionalmente muito pequenos, embora às vezes sejam encontrados restos de um terceiro dedo vestigial.[11][19]

Os tiranossaurídeos andavam exclusivamente sobre os membros posteriores, de modo que os ossos das pernas eram enormes. Em contraste com os membros anteriores, os membros posteriores eram mais longos em comparação com o tamanho do corpo do que quase todos os outros terópodes. Os juvenis e até mesmo alguns adultos menores, como os tiranossauroides mais basais, tinham tíbias mais longas que os fêmures, uma característica de dinossauros cursoriais (de corrida rápida) como os ornitomimídeos. Adultos maiores tinham proporções de pernas características de animais mais lentos, mas não na extensão vista em outros grandes terópodes como abelissaurídeos ou carnossauros. Os terceiros metatarsos dos tiranossaurídeos foram comprimidos entre o segundo e o quarto metatarsos, formando uma estrutura conhecida como arctometatarso.[11]
Não está claro quando o arctometatarso evoluiu pela primeira vez; não estava presente nos primeiros tiranossauróides como Dilong,[20] mas foi encontrado no Appalachiosaurus posterior.[15] Essa estrutura também caracterizou troodontídeos, ornitomimídeos e caenagnatídeos,[21] mas sua ausência nos primeiros tiranossauróides indica que ela foi adquirida por evolução convergente.[20]
Classificação
editarO nome Deinodontidae foi cunhado por Edward Drinker Cope em 1866 para esta família,[22] e continuou a ser usado no lugar do nome mais recente Tyrannosauridae até a década de 1960.[23] O gênero tipo de Deinodontidae é Deinodon, que recebeu o nome de dentes isolados de Montana.[24] No entanto, em uma revisão de tiranossaurídeos norte-americanos em 1970, Dale Russell concluiu que Deinodon não era um táxon válido e usou o nome Tyrannosauridae no lugar de Deinodontidae, afirmando que isso estava de acordo com as regras do ICZN.[13] Portanto, Tyrannosauridae é preferido por especialistas modernos.[6]
O tiranossauro foi nomeado por Henry Fairfield Osborn em 1905, junto com a família Tyrannosauridae.[25] O nome é derivado das palavras gregas antigas τυραννος (tyrannos) ('tirano') e σαυρος (sauros) ('lagarto'). O sufixo muito comum -idae é normalmente anexado a nomes de famílias zoológicas e é derivado do sufixo grego -ιδαι -idai, que indica um substantivo plural.[26]
Taxonomia
editarTyrannosauridae é uma família na taxonomia Linnaeana baseada em classificação, dentro da superfamília Tyrannosauroidea e da subordem Theropoda.[11]

Tyrannosauridae é incontroversamente dividido em duas subfamílias. Albertosaurinae compreende os gêneros norte-americanos Albertosaurus e Gorgosaurus, enquanto Tyrannosaurinae inclui Daspletosaurus, Teratophoneus, Bistahieversor, Tarbosaurus, Nanuqsaurus, Zhuchengtyrannus e o próprio Tyrannosaurus.[27] Alguns autores incluem as espécies Gorgosaurus libratus no gênero Albertosaurus e Tarbosaurus bataar no gênero Tyrannosaurus,[15][6][28] enquanto outros preferem manter Gorgosaurus e Tarbosaurus como gêneros separados.[11][12] Membro do clado Albertosaurinae são caracterizados por estruturas mais delgadas, crânios mais baixos e tíbias proporcionalmente mais longas do que os tiranossaurinos.[11] Nos membros de Tyrannosaurinae, a crista sagital nos parietais continua para frente nos frontais.[12] Em 2014, Lü Junchang et al. descreveu o Alioramini como uma tribo dentro dos Tyrannosauridae contendo os gêneros Alioramus e Qianzhousaurus. Sua análise filogenética indicou que a tribo estava localizada na base dos Tyrannosaurinae.[29][30] Alguns autores, como George Olshevsky e Tracy Ford, criaram outras subdivisões ou tribos para várias combinações de tiranossaurídeos dentro das subfamílias.[31][32] No entanto, eles não foram definidos filogeneticamente e geralmente consistiam em gêneros que agora são considerados sinônimos de outros gêneros ou espécies.[17]
Subfamílias adicionais foram nomeadas para gêneros mais fragmentários, incluindo Aublysodontinae e Deinodontinae. No entanto, os gêneros Aublysodon e Deinodon são geralmente considerados duvidosos, então eles e suas subfamílias epônimas são geralmente excluídos das taxonomias de tiranossaurídeos. Um tiranossaurídeo adicional, Raptorex, foi inicialmente descrito como um tiranossauroide mais primitivo, mas provavelmente representa um tiranossaurídeo jovem semelhante ao Tarbosaurus. No entanto, como é conhecido apenas em uma amostra juvenil, também é atualmente considerado um nomen dubium.[33]
Filogenia
editarUm estudo relacionado também observou um mecanismo de travamento na mandíbula inferior compartilhado entre os dois gêneros.[34] Em um artigo separado, Currie observou a possibilidade de que Alioramus pode representar um Tarbosaurus juvenil, mas afirmou que a contagem de dentes muito maior e cristas nasais mais proeminentes em Alioramus sugerem que é um gênero distinto. Da mesma forma, Currie usa a alta contagem de dentes do Nanotyrannus para sugerir que ele pode ser um gênero distinto,[12] em vez de um Tiranossauro juvenil, como muitos outros especialistas acreditam.[11][35] No entanto, a descoberta e a descrição do Qianzhousaurus revelam que Alioramus não é um parente próximo do Tarbosaurus, em vez disso, pertence a uma tribo recentemente descrita de tiranossaurídeos; o Alioramini. Qianzhousaurus revela ainda que semelhantes tiranossaurídeos de focinho longo foram amplamente distribuídos por toda a Ásia e teriam compartilhado o mesmo ambiente, evitando a competição com tiranossaurídeos maiores e mais robustos ao caçar presas diferentes.[36]
O cladograma abaixo é de Fiorillo e sua equipe, montado em 2014.[27]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Loewen e sua equipe fizeram este cladograma em 2013[37]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
editarCrescimento
editarO paleontólogo Gregory Erickson e seus colegas estudaram o crescimento e a história de vida dos tiranossaurídeos. A análise da histologia óssea pode determinar a idade de um espécime quando morreu. As taxas de crescimento podem ser examinadas quando a idade de vários indivíduos é plotada em relação ao seu tamanho em um gráfico. Erickson mostrou que, depois de muito tempo como juvenis, os tiranossaurídeos passaram por enormes surtos de crescimento por cerca de quatro anos no meio de suas vidas. Depois que a fase de crescimento rápido terminou com a maturidade sexual, o crescimento diminuiu consideravelmente em animais adultos. Uma curva de crescimento dos tiranossaurídeos tem formato de S, com a taxa máxima de crescimento dos indivíduos por volta dos 14 anos de idade.[38]

Estima-se que o menor Tyrannosaurus rex conhecido (LACM 28471, o "terópode da Jordânia") pesava apenas 29,9 kg com apenas 2 anos de idade, enquanto o maior, como FMNH PR2081 ("Sue"), provavelmente pesava cerca de 5.654 kg, estimado em 28 anos, uma idade que pode ter sido próxima do máximo para a espécie.[38] Os juvenis de T. rex permaneceram abaixo de 1.800 kg até aproximadamente 14 anos de idade, quando o tamanho do corpo começou a aumentar drasticamente. Durante esta fase de crescimento rápido, um jovem T. rex ganharia uma média de 600 kg por ano nos próximos quatro anos. Isso diminuiu após 16 anos e, aos 18 anos de idade, a curva se estabiliza novamente, indicando que o crescimento diminuiu drasticamente.[39] Por exemplo, apenas 600 kg separaram o "Sue" de 28 anos de um espécime canadense de 22 anos (RTMP 81.12.1).[38] Essa mudança repentina na taxa de crescimento pode indicar maturidade física, uma hipótese que é apoiada pela descoberta de tecido medular no fêmur de um T. rex de 18 anos de Montana (MOR 1125, também conhecido como "B-rex") .[40] O tecido medular é encontrado apenas em aves fêmeas durante a ovulação, indicando que o "B-rex" estava em idade reprodutiva.[41]
Outros tiranossaurídeos exibem curvas de crescimento extremamente semelhantes, embora com taxas de crescimento mais baixas correspondendo a seus tamanhos adultos mais baixos.[42] Comparado aos membros de Albertosaurinae, o Daspletosaurus mostrou uma taxa de crescimento mais rápida durante o período de crescimento rápido devido ao seu maior peso adulto. A taxa máxima de crescimento em Daspletosaurus foi de 180 kg por ano, com base em uma estimativa de massa de 1.800 kg em adultos. Outros autores sugeriram maiores pesos adultos para Daspletosaurus; isso mudaria a magnitude da taxa de crescimento, mas não o padrão geral.[38] O Albertosaurus mais jovem conhecido é um de dois anos descoberto no leito ósseo da Ilha Seca, que pesava cerca de 50 kg e media pouco mais de 2 metros de comprimento. O espécime de 10 metros da mesma pedreira é o mais antigo e maior conhecido, com 28 anos de idade. Estima-se que a taxa de crescimento mais rápida ocorra em torno de 12 a 16 anos de idade, atingindo 122 kg por ano, com base em um adulto de 1.300 kg, que é cerca de um quinto da taxa de T.-rex. Para Gorgosaurus, a taxa de crescimento máxima calculada é de cerca de 110 kg durante a fase de crescimento rápido, que é comparável à do Albertosaurus.[38]
A descoberta de um tiranossauro embrionário de um gênero ainda desconhecido sugere que os tiranossaurídeos desenvolveram suas características esqueléticas distintas enquanto se desenvolviam no ovo. Além disso, o tamanho do espécime, um dentário de 2,8 cm do maxilar inferior encontrado na Formação Two Medicine de Montana em 1983 e uma garra do pé encontrada na Formação Horseshoe Canyon em 2018 e descrita em 2020, sugere que o recém-nascido os tiranossaurídeos nasceram com crânios do tamanho de um camundongo ou roedores de tamanho semelhante e podem ter sido aproximadamente do tamanho de um cachorro pequeno ao nascer. Acredita-se que o espécime da mandíbula tenha vindo de um animal de aproximadamente 2,5 pés, enquanto acredita-se que a garra pertença a um espécime medindo cerca de 3 pés. Embora cascas de ovos não tenham sido encontradas em associação com nenhum dos espécimes, o local onde esses tiranossaurídeos recém-nascidos foram descobertos sugere que esses animais estavam usando os mesmos locais de ninho que outras espécies com as quais viviam e predavam.[43] A falta de cascas de ovos associadas a esses espécimes também abriu especulações para a possibilidade de que os tiranossaurídeos colocassem ovos de casca mole, como acredita-se que os gêneros Mussaurus e Protoceratops tenham feito.[44]
Pegadas fósseis da Formação Wapiti sugerem que, à medida que os tiranossaurídeos cresciam, os pés se tornavam mais largos com dedos mais grossos para suportar seu peso. Os pés mais largos sugerem que os tiranossaurídeos adultos eram mais lentos do que seus descendentes.[45][46]
História de vida
editar
O fim da fase de crescimento rápido sugere o início da maturidade sexual no Albertosaurus, embora o crescimento tenha continuado a um ritmo mais lento ao longo da vida dos animais.[38][42] A maturação sexual enquanto ainda cresce ativamente parece ser uma característica compartilhada entre dinossauros pequenos[47] e grandes[48], bem como em grandes mamíferos, como humanos e elefantes.[48] Esse padrão de maturação sexual relativamente precoce difere notavelmente do padrão das aves, que atrasam sua maturidade sexual até que tenham terminado de crescer.[48][49]
Ao tabular o número de espécimes de cada faixa etária, Erickson e seus colegas conseguiram tirar conclusões sobre a história de vida em populações de tiranossaurídeos. Sua análise mostrou que, enquanto os juvenis eram raros no registro fóssil, os subadultos na fase de crescimento rápido e os adultos eram muito mais comuns. Mais da metade dos espécimes de T. rex conhecidos parecem ter morrido dentro de seis anos após atingirem a maturidade sexual, um padrão que também é visto em outros tiranossaurídeos e em algumas aves e mamíferos grandes e de vida longa hoje. Essas espécies são caracterizadas por altas taxas de mortalidade infantil, seguidas por uma mortalidade relativamente baixa entre os juvenis. A mortalidade aumenta novamente após a maturidade sexual, em parte devido ao estresse da reprodução. Embora isso possa ser devido a vieses de preservação ou coleta, Erickson levantou a hipótese de que a diferença se devia à baixa mortalidade entre os juvenis acima de um determinado tamanho, o que também é visto em alguns grandes mamíferos modernos, como elefantes. Essa baixa mortalidade pode ter resultado da falta de predação, já que os tiranossaurídeos superaram todos os predadores contemporâneos em tamanho aos dois anos de idade. Os paleontólogos não encontraram restos de Daspletossauro suficientes para uma análise semelhante, mas Erickson observa que a mesma tendência geral parece se aplicar.[42]
Os tiranossaurídeos passaram metade de sua vida na fase juvenil antes de atingir o tamanho quase máximo em apenas alguns anos.[38] Isso, juntamente com a completa falta de predadores de tamanho intermediário entre os enormes tiranossaurídeos adultos e outros pequenos terópodes, sugere que esses nichos podem ter sido preenchidos por tiranossaurídeos juvenis. Isso é visto em dragões-de-komodo modernos, onde os filhotes começam como insetívoros que vivem em árvores e amadurecem lentamente em enormes predadores capazes de derrubar grandes vertebrados.[11] Por exemplo, Albertosaurus foram encontrados em agregações que alguns sugeriram para representar bandos, com indíviduos de idades mistas.[50][51]
Paleogeografia
editar
Enquanto os primevos tiranossauróides são encontrados em todos os três continentes do norte, os fósseis de tiranossaurídeos são conhecidos apenas da América do Norte e da Ásia. Às vezes, restos fragmentários descobertos no Hemisfério Sul foram relatados como "tiranossaurídeos do Hemisfério Sul", embora estes pareçam ter sido fósseis de abelissaurídeos erroneamente identificados.[52] O momento exato e o local de origem da família permanecem desconhecidos devido ao pobre registro fóssil na parte média do Cretáceo em ambos os continentes, embora os primeiros tiranossaurídeos confirmados tenham vivido no início do estágio Campaniano no oeste da América do Norte.[11]
Restos de tiranossaurídeos nunca foram recuperados do leste da América do Norte, enquanto tiranossaurídeos mais basais, como Dryptosaurus e Appalachiosaurus, persistiram lá até o final do Cretáceo, indicando que estes devem ter evoluído ou se dispersado no oeste da América do Norte depois que o continente foi dividido ao meio pelo Mar Interior Ocidental no meio do Cretáceo.[15] Fósseis de tiranossaurídeos foram encontrados no Alasca, o que pode ter fornecido uma rota de dispersão entre a América do Norte e a Ásia.[53] Alioramus e Tarbosaurus são relacionados em uma análise cladística, formando um ramo asiático único da família.[17] Isso foi posteriormente refutado com a descoberta do Qianzhousaurus e a descrição da tribo Alioramini. Dentes de tiranossaurídeos de uma grande espécie de variedade desconhecida foram descobertos na Península de Nagasaki por pesquisadores do Museu de Dinossauros da Prefeitura de Fukui, expandindo ainda mais o alcance do grupo. Os dentes foram estimados em 81 milhões de anos (estágio Campaniano).[54]
Das duas subfamílias, Tyrannosaurinae parece ter sido mais difundida. Albertosaurinae são desconhecidos na Ásia, que foi o lar de tiranossaurinos, como Tarbosaurus e Zhuchengtyrannus, e Qianzhousaurus e Alioramus dos Alioramini. Ambas as subfamílias Tyrannosaurinae e Albertosaurinae estavam presentes nos estágios Campaniano e Maastrichtiano inicial da América do Norte, com tiranossaurídeos como Daspletosaurus variando por todo o Interior Ocidental, enquanto os abertossaurinos Albertosaurus e Gorgosaurus são atualmente conhecidos apenas na parte noroeste do continente.[55]
No final do estágio Maastrichtiano, os poucos membros de Albertosaurinae parecem ter sido extintos, enquanto o maior representente de Tyrannosaurinae, Tyrannosaurus, vagava de Saskatchewan ao Texas. Este padrão é espelhado em outros táxons de dinossauros norte-americanos. Durante o Campaniano e início do Maastrichtiano, hadrossaurídeos como lambeossauros e ceratopsianos centrosaurinos são comuns no noroeste, enquanto hadrossaurídeos e chasmosaurinos eram mais comuns no sul. No final do Cretáceo, os centrosaurinos são desconhecidos e os lambeossaurídeos são raros, enquanto os hadrossaurídeos e os chasmosaurinos eram comuns em todo o Interior Ocidental.[11] Um estudo publicado na revista Scientific Reports em 2 de fevereiro de 2016, por Steve Brusatte, Thomas Carr et al. indica que durante o Maastrichtiano posterior, o próprio Tyrannosaurus pode ter sido parcialmente responsável pela extinção dos outros tiranossaurídeos na maior parte do oeste da América do Norte. O estudo indica que este pode ter sido um imigrante da Ásia em oposição a ter evoluído na América do Norte (possivelmente um descendente do Tarbosaurus intimamente relacionado) que suplantou e superou outros tiranossaurídeos. Esta teoria é ainda apoiada pelo fato de que poucos ou nenhum outro tipo de tinossaurídeos são encontrados dentro do alcance conhecido do T. rex.[56]
Linha do tempo dos gêneros
editar
Ver também
editarReferências
- ↑ Holtz, Thomas R. Jr. (2012) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix. Arquivado em 2017-08-12 no Wayback Machine
- ↑ «Conheça 4 curiosidades sobre os tiranossauros». Galileu. 8 de abril de 2019. Consultado em 10 de outubro de 2022
- ↑ a b c Hutchinson, John R.; Bates, Karl T.; Molnar, Julia; Allen, Vivian; Makovicky, Peter J.; Claessens, Leon (2011). «A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth». PLOS ONE (em inglês). 6 (10): e26037. Bibcode:2011PLoSO...626037H. PMC 3192160
 . PMID 22022500. doi:10.1371/jornal.pone.0026037
. PMID 22022500. doi:10.1371/jornal.pone.0026037
- ↑ Therrien, F.; Henderson, D. M. (2007). «My theropod is bigger than yours ... or not: estimating body size from skull length in theropods». jornal of Vertebrate Paleontology. 27 (1): 108–115. ISSN 0272-4634. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2
- ↑ Hartman, Scott (7 de julho de 2013). «Mass estimates: North vs South redux» (em inglês). Scott Hartman's Skeletal Drawing.com. Consultado em 24 de agosto de 2013
- ↑ a b c d e Carpenter, Ken. (1992). «Tyrannosaurids (Dinosauria) of Asia and North America». In: Mateer, Niall J.; Chen Peiji. Aspects of Nonmarine Cretaceous Geology (em inglês). Beijing: China Ocean Press. pp. 250–268
- ↑ Breithaupt, B.H.; Southwell, E.H.; Matthews, N.A. (18 de outubro de 2005). «In Celebration of 100 anos of Tyrannosaurus rex: Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the Earliest Discoveries of Tyrannosaurus rex in the West». Abstracts with Programs. 2005 Salt Lake City Annual Meeting (em inglês). 37. Geological Society of America. 406 páginas. Consultado em 8 outubro de 2008
- ↑ Breithaup, BH; Southwell EH; Matthews NA (2006). «Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West». New Mexico Museum of Natural History and Science Bulletin (em inglês). 35: 257–258
- ↑ Bakker R.T; Williams M.; Currie P. (1988). «Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana». Hunteria (em inglês). 1: 1–30
- ↑ a b Kurzanov, Sergei M. «A new carnosaur from the Late Cretaceous of Nogon-Tsav, Mongolia». The Joint Soviet-Mongolian Paleontological Expedition Transactions (em russo). 3: 93–104
- ↑ a b c d e f g h i j k l Holtz, Thomas R. (2004). «Tyrannosauroidea». In: Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria Second ed. Berkeley: University of California Press. pp. 111–136. ISBN 978-0-520-24209-8
- ↑ a b c d e Currie, Philip J. (2003). «Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta» (PDF). Acta Palaeontologica Polonica (em inglês). 48 (2): 191–226
- ↑ a b Russell, Dale A. (1970). «Tyrannosaurs from the Late Cretaceous of western Canada». National Museum of Natural Sciences Publications in Paleontology. 1: 1–34
- ↑ Maleev, Evgeny A. (1955). «New carnivorous dinosaurs from the Upper Cretaceous of Mongolia» (PDF). Doklady Akademii Nauk SSSR (em russo). 104 (5): 779–783
- ↑ a b c d e Carr, Thomas D.; Williamson, Thomas E.; Schwimmer, David R. (2005). «A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama». jornal of Vertebrate Paleontology (em inglês). 25 (1): 119–143. ISSN 0272-4634. doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2
- ↑ Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (30 de março de 2017). «A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system». Scientific Reports (em inglês). 7 (1): 44942. Bibcode:2017NatSR...744942C. ISSN 2045-2322. PMC 5372470 . PMID 28358353. doi:10.1038/srep44942
- ↑ a b c Currie, Philip J.; Hurum, Jørn H; Sabath, Karol. (2003). «Skull structure and evolution in tyrannosaurid phylogeny» (PDF). Acta Palaeontologica Polonica. 48 (2): 227–234
- ↑ a b c d e f g Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84–89. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ↑ Quinlan, Elizabeth D.; Derstler, Kraig; Miller, Mercedes M. (2007). «Anatomy and function of digit III of the Tyrannosaurus rex manus». Geological Society of America Annual Meeting — Abstracts with Programs: 77. Consultado em 15 de janeiro de 2008. Arquivado do original em 24 de fevereiro de 2008 [Apenas abstract]
- ↑ a b Xu Xing, X; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; Jia Chengkai. (2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature. 431 (7009): 680–684. Bibcode:2004Natur.431..680X. PMID 15470426. doi:10.1038/nature02855
- ↑ Holtz, Thomas R. (1994). «The phylogenetic position of the Tyrannosauridae: implications for theropod systematics». Journal of Paleontology. 68 (5): 1100–1117. JSTOR 1306180. doi:10.1017/S0022336000026706
- ↑ Cope E.D. (1866). «Discovery of a gigantic dinosaur in the Cretaceous of New Jersey». Proceedings of the Academy of Natural Sciences of Philadelphia. 18: 275–279
- ↑ Maleev E.A. (1955). «New carnivorous dinosaurs from the Upper Cretaceous Mongolia». Doklady Akademii Nauk SSSR. 104 (5): 779–783
- ↑ Leidy, Joseph (1856). «Notice of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the badlands of the Judith River, Nebraska Territory». Proceedings of the Academy of Natural Sciences of Philadelphia. 8: 72–73
- ↑ Osborn, Henry F. (1905). «Tyrannosaurus and other Cretaceous carnivorous dinosaurs». Bulletin of the American Museum of Natural History. 21 (3): 259–265. doi:10.1111/j.1468-5965.2007.00735_17.x. hdl:2246/1464
- ↑ Liddell, Henry G.; Scott, Robert (1980). Greek-English Lexicon Abridged ed. Oxford: Oxford University Press. ISBN 978-0-19-910207-5
- ↑ a b Fiorillo, A. R.; Tykoski, R. S. (2014). Dodson, Pete, ed. «A Diminutive New Tyrannosaur from the Top of the World». PLoS ONE (em inglês). 9 (3): e91287. Bibcode:2014PLoSO...991287F. PMC 3951350 . PMID 24621577. doi:10.1371/journal.pone.0091287
- ↑ Paul, Gregory S. (1988). Predatory Dinosaurs of the World. Nova Iorque: Simon & Schuster. pp. 464pp. ISBN 978-0-671-61946-6
- ↑ Lü, Junchang; Yi, Laiping; Brusatte, Stephen L.; Yang, Ling; Li, Hua; Chen, Liu (2014). «A new clade of Asian Late Cretaceous long-snouted tyrannosaurids». Nature Communications. 5: 3788. Bibcode:2014NatCo...5.3788L. PMID 24807588. doi:10.1038/ncomms4788
- ↑ «Pinocchio rex long-snouted tyrannosaur discovered in Asia». 7 de maio de 2014
- ↑ Olshevsky, George (1995). «The origin and evolution of the tyrannosaurids». Kyoryugaku Saizensen [Dino Frontline]. 9–10: 92–119
- ↑ Olshevsky G.; Ford T.L. (1995). «The origin and evolution of the Tyrannosauridae, part 2 [in Japanese]». Dino Frontline. 6: 75–99
- ↑ Fowler, DW; Woodward, HN; Freedman, EA; Larson, PL; Horner, JR (2011). «Reanalysis of "Raptorex kriegsteini": A Juvenile Tyrannosaurid Dinosaur from Mongolia». PLOS ONE. 6 (6): e21376. Bibcode:2011PLoSO...6E1376F. PMC 3126816 . PMID 21738646. doi:10.1371/journal.pone.0021376
- ↑ Hurum, Jørn H.; Sabath, Karol. (2003). «Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared» (abstract). Acta Palaeontologica Polonica. 48 (2): 161–190
- ↑ Carr, Thomas D. (1999). «Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)». Journal of Vertebrate Paleontology. 19 (3): 497–520. doi:10.1080/02724634.1999.10011161
- ↑ «Newly found dinosaur is long-nosed cousin of Tyrannosaurus rex». ScienceDaily
- ↑ Loewen, M.A.; Irmis, R.B.; Sertich, J.J.W.; Currie, P. J.; Sampson, S. D. (2013). Evans, David C, ed. «Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans». PLoS ONE. 8 (11): e79420. Bibcode:2013PLoSO...879420L. PMC 3819173 . PMID 24223179. doi:10.1371/journal.pone.0079420
- ↑ a b c d e f g Erickson, Gregory M., GM; Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; Brochu, Christopher A. (2004). «Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs». Nature. 430 (7001): 772–775. Bibcode:2004Natur.430..772E. PMID 15306807. doi:10.1038/nature02699.
Sampled longevities for T. rex ranged from 2 to 28 years
- ↑ Horner, J.R.; Padian, K. (2004). «Age and growth dynamics of Tyrannosaurus rex». Proceedings of the Royal Society of London B. 271 (1551): 1875–1880. PMC 1691809 . PMID 15347508. doi:10.1098/rspb.2004.2829 Verifique o valor de
|name-list-format=amp(ajuda) - ↑ Lee, Andrew H.; Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. PMC 2206579 . PMID 18195356. doi:10.1073/pnas.0708903105
- ↑ Schweitzer, M.H., Wittmeyer, J.L., & Horner, J.R. (2005). «Gender-specific reproductive tissue in ratites and Tyrannosaurus rex». Science. 308 (5727): 1456–1460. Bibcode:2005Sci...308.1456S. PMID 15933198. doi:10.1126/science.1112158. Consultado em 2 de dezembro de 2019. Cópia arquivada em 10 de novembro de 2021
- ↑ a b c Erickson, G.M., Currie, P.J., Inouye, B.D., & Winn, A.A. (2006). «Tyrannosaur life tables: an example of nonavian dinosaur population biology». Science. 313 (5784): 213–217. Bibcode:2006Sci...313..213E. PMID 16840697. doi:10.1126/science.1125721. Consultado em 2 de dezembro de 2019. Cópia arquivada em 10 de novembro de 2021
- ↑ «First tyrannosaur embryo fossils revealed». Science. 19 de outubro de 2020. Consultado em 23 de novembro de 2020. Cópia arquivada em 5 de dezembro de 2020
- ↑ October 2020, Laura Geggel-Associate Editor 15 (15 de outubro de 2020). «1st of their kind baby tyrannosaur fossils unearthed». livescience.com. Consultado em 17 de outubro de 2020. Cópia arquivada em 17 de outubro de 2020
- ↑ Enriquez, Nathan James; Campione, Nicolas; Bell, Phil. «Fat-footed tyrannosaur parents couldn't keep up with their skinnier offspring, fossil footprints reveal». The Conversation. Consultado em 28 de abril de 2021. Cópia arquivada em 26 de abril de 2021
- ↑ Enriquez, Nathan J.; Campione, Nicolás E.; Brougham, Tom; Fanti, Federico; White, Matt A.; Sissons, Robin L.; Sullivan, Corwin; Vavrek, Matthew J.; Bell, Phil R. (21 de abril de 2021). «Exploring possible ontogenetic trajectories in tyrannosaurids using tracks from the Wapiti Formation (upper Campanian) of Alberta, Canada». Journal of Vertebrate Paleontology. 40 (6): e1878201. doi:10.1080/02724634.2021.1878201
- ↑ Erickson, Gregory M.; Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark; Xu, Xing (2007). «Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition». Biology Letters. 3 (5): 558–561. PMC 2396186 . PMID 17638674. doi:10.1098/rsbl.2007.0254
- ↑ a b c Lee, Andrew H.; Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». PNAS. 105 (2): 582–587. Bibcode:2008PNAS..105..582L. PMC 2206579 . PMID 18195356. doi:10.1073/pnas.0708903105
- ↑ Ricklefs, Robert E. (2007). «Tyrannosaur ageing». Biology Letters. 3 (2): 214–217. PMC 2375931 . PMID 17284406. doi:10.1098/rsbl.2006.0597
- ↑ Tanke, Darren H.; Currie, Philip J. (1998). «Head-biting behavior in theropod dinosaurs: paleopathological evidence» (PDF). Gaia. 15: 167–184. Consultado em 13 de setembro de 2008. Cópia arquivada (PDF) em 27 de fevereiro de 2008 [not printed until 2000]
- ↑ Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; Murphy, Nate (2005). «An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)». In: Carpenter, Kenneth. The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 313–324. ISBN 978-0-253-34539-4
- ↑ Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A.; Gillette, David D.; Norell, Mark A.; Olshevsky, George; Parrish, J. Michael; Weishampel, David B. (1993). «Abelisaurus». Age of Dinosaurs. [S.l.]: Publications International, LTD. p. 105. ISBN 978-0-7853-0443-2
- ↑ Fiorillo, Anthony R.; Gangloff, Roland A. (2000). «Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on arctic dinosaur paleoecology». Journal of Vertebrate Paleontology. 20 (4): 675–682. ISSN 0272-4634. doi:10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2
- ↑ «These 81-million-year-old teeth are the first bit of evidence of Japan's largest ever dinosaur | Irish Examiner». Consultado em 10 de março de 2016. Cópia arquivada em 12 de abril de 2016
- ↑ Thomas R. Holtz Jr. (1994). «The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics». Journal of Paleontology. 68 (5): 1100–1117. JSTOR 1306180. doi:10.1017/S0022336000026706
- ↑ Geggel, Laura (29 de fevereiro de 2016). «T. Rex Was Likely an Invasive Species». livescience.com. Consultado em 5 de janeiro de 2021. Cópia arquivada em 14 de julho de 2019
Ligações externas
editar Media relacionados com Tyrannosauridae no Wikimedia Commons
Media relacionados com Tyrannosauridae no Wikimedia Commons O Wikispecies possui informações sobre: Tyrannosauridae
O Wikispecies possui informações sobre: Tyrannosauridae